| <<Vol. 2 Table of Contents | |
|
|
|
|
2007 |
Volume 2, pp 1-25 |

Tool-related Cognition in New Caledonian Crows
Lucas A. Bluff, Alex A. S. Weir, Christian Rutz, Joanna H. Wimpenny, and Alex Kacelnik
University of Oxford
|
|
|||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||
Our group has studied tool-oriented behaviour (TOB) in New Caledonian crows (Corvus moneduloides) for the past six years (Figure 1). This followed the first systematic documentation of wild New Caledonian crow behaviour by Gavin Hunt and associates from Auckland University (Hunt, 1996, 2000a; Hunt, Corballis, & Gray, 2001; Hunt & Gray, 2003). Here we present a progress report that focuses on experimental studies with captive crows in Oxford. We discuss the results of our experiments with reference both to the field data collected by our New Zealand-based colleagues and to theoretical issues faced by other researchers in comparative cognition. We address—but do not solve—the epistemological difficulties inherent in attempts to uncover aspects of physical understanding and reasoning in non-human species. Our broad perspective is that while tool-related behaviour is not necessarily associated with unusually sophisticated cognition, it is likely to be unusually revealing about the cognitive processes and the level of understanding involved in animals’ manipulation of physical objects. Uniquely amongst non-mammals, New Caledonian crows make and use a number of distinct tool types in the wild and in captivity (Figure 2), some of which involve considerable processing of the raw materials (Hunt, 1996; Hunt & Gray, 2004a, 2004b). Tool use and manufacture by animals has attracted considerable interest from scientists and the general public, probably because of its rarity and its apparent association with the human lineage. Whilst it is now widely accepted that tool use per se is not indicative of unusual intelligence (e.g., Alcock, 1972; Beck, 1980, 1986; Hall, 1963; Hansell, 1987), the remarkable complexity of New Caledonian crows’ natural TOB has often led observers (lay people as well as scholars) to assume that this species may possess exceptionally advanced cognitive abilities. However, such first impressions should be treated as working hypotheses at best, as the motor complexity of a behaviour offers no guide to its underlying cognitive complexity: There are many examples of complex architecture—typically in the form of nests—throughout the animal kingdom (Hansell, 2005), none of which are thought to require generally elevated cognitive abilities.
Natural behaviour arises through the interaction of the genetic endowment with a number of ontogenetic processes that include trial-and-error, associative and social learning, and possibly reasoning. It is axiomatic that the contribution of each of these, and especially of ‘higher’ cognitive processes such as reasoning or insight, cannot be reliably inferred from observing spontaneous behaviour of wild animals without experimental investigation. Even when experimentation is possible, these processes are fiendishly elusive because virtually everything an adult animal does is affected by previous associative learning and generalisation. This difficulty is, of course, the strongest justification for the behaviourists’ reluctance to even invoke the possibility of entities such as reasoning or understanding, and we do have sympathy with this argument by parsimony. However, the complete learning history of anything but newborn individuals is never fully known. Therefore, although attributing innovative problem-solving to a hypothetical combination of reinforced learning and generalisation appears parsimonious because the mechanisms are uncontroversial, it is in fact just as speculative as accounts involving abstract forms of information processing that would qualify as ‘reasoning’ or ‘understanding’. We are fully aware that such information-processing mechanisms are not alternatives to associative learning but may act in concert to produce innovative behaviour. However, it would seem agreeable to most that, when observing a behavioural innovation, the more contrived the required generalisation from previously reinforced behaviour, the more plausible it is to invoke the existence of alternative mechanisms involving higher level concepts (we have made similar points elsewhere; see Weir & Kacelnik, 2006). This is the theoretical framework in which we seek to explore the involvement of mechanisms best defined in terms of their folk-psychological labels, such as reasoning, understanding, and creativity, in tool-oriented behaviour of our model species. Although TOB is not necessarily associated with unusual cognition, the making and using of tools may be especially revealing from a cognitive perspective. Making and using objects to act on other objects, such as food, may give us a valuable glimpse of an organism’s use of abstraction or generalisation in the physical domain. Tool use entails a behavioural richness that can be exploited in controlled experiments where it is unlikely that the subjects can ‘solve’ the task by deployment of a previously reinforced behaviour or a combination of such behaviours. It is challenging to design and interpret such experiments, but TOB offers a context in which the opportunity for innovation is greater than in situations where subjects are intensively trained to collect rewards using binary manipulanda such as pecking keys or levers. So far, we have investigated two different aspects of New Caledonian crows’ TOB: the epigenetic processes that lead to the development of fully-functional TOB, and the cognitive processes that are involved in the deployment of TOB in adult crows. While both levels of enquiry are necessary to understand this species’ cognitive capacities, it is important to recognise that, at different periods within a crow’s life, different mechanisms may govern the expression of functional tool use. For example, observing the spontaneous emergence of a non-reinforced behaviour during early development leads us to infer the presence of a genetic propensity for that behaviour, but this does not imply that its subsequent deployment is unaccompanied by information-processing mechanisms that would qualify as higher cognition. Indeed, genetic predispositions are thought to be the building blocks through which human children learn about the properties of objects (Lockman, 2000), yet older humans are able to devise solutions to novel problems through processes such as reasoning. We acknowledge that exploring the cognition behind action, especially in the context of physical understanding, presents major epistemological challenges, mostly because of the difficulty in isolating situations that admit a single cognitive account. In fact, it is inherently difficult even to define what it means to say that someone ‘understands’ their actions: A recent philosophical review concluded that humans describe themselves as ‘understanding’ something if they believe that they are not hampered in reaching a relevant, currently active goal by lack of knowledge about that ‘something’ (Overskeid, 2005). Applying such a definition based on goals and knowledge to non-humans is not easy. Most research to date into the causal understanding of non-humans has been framed in terms of whether the subjects’ behaviour is generated by associative versus inferential (or reasoning) processes (e.g., Call, 2004; Dickinson, 2001), where the ‘understanding’ of the causal basis of something (an event or action) is equated with the ability to infer the causal agent. However, this debate has been hampered by a lack of precision in the specification of what is meant by reasoning or inferring: As Penn and Povinelli (2007) point out, the advocates for such processes tend to formulate their arguments verbally, rather than mathematically, which makes them almost impossible to refute. Some authors do explicitly define higher-order reasoning—for example, as "processes [that] can be characterized as reflective […], rule-based […], and deliberate […] and [which] operate on conscious propositional knowledge in a controlled (i.e., slow, effortful, conscious, and/or intentional) manner" (de Houwer, Beckers, & Vandorpe, 2005, p. 240). However, most researchers (including, at times, ourselves in previous publications) informally invoke reasoning or other cognitive processes for situations where associative learning seems insufficient to explain the subjects’ behaviour. This has led to a technical debate about the mathematical properties of associative learning and what behaviours it can and cannot explain (e.g., Dickinson, 2001) and whether other algorithms such as causal Bayes nets provide a better explanation (e.g., Blaisdell, Sawa, Leising, & Waldmann, 2006). We feel that this debate does not address one of the most interesting questions: Are any non-human animals capable of understanding and reasoning about causality in a qualitatively similar way to humans? Penn and Povinelli point out that human causal knowledge is ‘theory-like,’ "in the sense that it provides principled, allocentric, coherent, abstract explanations for the unobservable causal mechanisms that govern a given domain," and suggest that a salient feature of these theories "is that they can be generalized freely to disparate concrete examples that share little to no perceptually based featural similarity" (Penn & Povinelli, 2007, p.107). Mechanisms such as associative learning are undoubtedly involved in the process of forming these theories, but the key point is that behaviour should be influenced by the individual’s understanding, rather than purely by the subject’s previous reinforcement history. The main goal in our laboratory work with New Caledonian crows is to establish whether ‘theory-like’ causal knowledge underlies their TOB—or in other words, whether they understand how their tools work. In spite of all the caveats listed above, we still feel that it is helpful to distinguish between behaviour shaped by known or easily conceived experiences of reinforcement (i.e., trial-and-error learning) and behaviour which appears to result from an abstract process of inference. It is worth noting that the methods typically used to investigate cognition in non-verbal animals are fundamentally different from those employed in studies with humans. To probe adult humans’ understanding of the physics underlying their use of a tool, one can ask them to verbalise their reasoning. This does not prove that their actions are guided by their expressed beliefs (which could be true or false regarding real physics), but it may tell us if the subjects feel that they understand why or how the tool works (Overskeid, 2005). Regardless of whether the use of the tool is competent, and regardless of whether the theory expressed to justify actions is correct, one can infer something about the existence and quality of understanding (a narrative based on a superstitious belief such as the gamblers’ fallacy is a mistaken but interesting form of understanding). Even this limited level of enquiry is impossible with non-human animals, so we have to rely on observing and interpreting tool-using performance and to focus on definitions of understanding that presuppose true beliefs. We are condemned to being over-conservative and denying any understanding to an animal that does have a theory and guides its behaviour by it, if this theory happens to be wrong. Our general experimental approach is to present our subjects with problems that, while being within the range of situations that members of the species are inclined to tackle, are as novel (namely as different from previous experiences in which reinforcement for behavioural components have occurred) as possible (similar to the approach taken by Hauser, 1997; Povinelli, 2000; Visalberghi, 1997). If subjects possessed abstract, theory-like explanations for how and why their tools operate, we could expect that in most such experiments they would solve the problems within the first few trials. In contrast, if their behavioural modification was governed by reinforcement, on most tasks most subjects would not be immediately successful. Each single experiment, therefore, provides limited information regarding how much of the observed behaviour is strictly novel, the outcome of reasoning, or both, particularly since (a) no task in which the birds willingly participate can ever be absolutely novel, (b) problems do not remain novel after the first trial, and (c) adult subjects may always generalise to some extent from previous experience. This means that, even though we transform the tasks as radically as possible from one experiment to another, there is no discrete threshold separating solutions produced by pure reinforcement of random behaviour from those resulting from understanding or reasoning. In summary, it is very difficult to define concepts that are used intuitively in every-day language to denote high-level cognition, let alone to investigate them empirically in animals. Reasoning, understanding, and logical inference are neither the opposite of genetically-channelled actions (which do not require reinforcement), nor of associatively learnt actions (which do). While parsimony advises against invoking these very opaque processes when more transparent ones are available, we still believe that the richer the number of task transformations that subjects can tolerate and solve, the more useful it is to invoke them. The main part of this review consists of two broad sections distinguishing the development and deployment of TOB. For some of our most cherished questions, such as the extent to which New Caledonian crows can use reasoning to solve physical problems, or what ecological and evolutionary circumstances led to the unusual behaviour of this species, we can only offer informed speculation. However, we do hope to show that some parts of the puzzle are being unravelled and that there is good reason to be optimistic about future progress, provided we advance with caution and are able to proceed simultaneously with experimental and observational research. Tool-oriented Behaviour in the Wild
The archipelago of New Caledonia is situated some 1,500 km east of the Australian mainland. New Caledonian crows are endemic to the main island of Grande Terre and one of its sister islands, Maré, where they were introduced by humans (ca. 1850s; Délacour, 1966). Crow tool use is featured in New Caledonian folklore and early European accounts, and stick tool use was reported by Orenstein in 1972. However, it was not until 1996 that the complexity and diversity of New Caledonian crow TOB was described by Gavin Hunt (Hunt, 1996). While many species use tools (Beck, 1980), the habitual use of multiple types of tool in the wild was thought to be restricted to primates (e.g., Parker & Gibson, 1977). Hunt and colleagues have demonstrated that, as a species, wild crows use a variety of tools, which they classified into three categories: straight sticks or leaf stems (Hunt, 2000b; Hunt & Gray, 2002; Rutledge & Hunt, 2004), hooked twigs or vines (Hunt, 1996; Hunt & Gray, 2004a), and tools torn from the leaves of the Pandanus tree (Figure 3; Hunt, 1996, 2000a; Hunt et al., 2001; Hunt, Corballis, & Gray, 2006; Hunt & Gray, 2003, 2004b). Like some other corvids (Cristol & Switzer, 1999), New Caledonian crows drop nuts to break them open (Layard & Layard, 1882), but using an unusual technique they wedge the nuts in tree forks before rolling them off (Hunt, Sakuma, & Shibata, 2002), perhaps to increase accuracy or efficacy of dropping. The shape of tools the crows produce from Pandanus leaves varies across New Caledonia, leading to the suggestion that tool-manufacturing skills may be transmitted and maintained culturally, perhaps even with a human-like ratchet effect where innovations are accumulated over time (Hunt, 2000a; Hunt & Gray, 2003). Since these data are observational, rather than experimental, and are based on the collection of tool counterparts made by an unknown number of individuals, the intriguing possibility of cultural transmission remains an untested hypothesis (see Laland & Janik, 2006). One of the main difficulties in resolving this issue is that it is not yet possible to compare variation in tool shapes produced by individuals from different populations, which requires the observation of large samples of individually-identifiable birds. Very little is currently known about the behavioural and social ecology of these crows in the wild, other than that they are social, that juveniles remain in some proximity to their parents for relatively long periods, and that they are found in dry and humid forest habitats in New Caledonia and in savannah and agricultural areas (reviewed in Kenward, Rutz, Weir, Chappell, & Kacelnik, 2004). Field studies with marked individuals by both research groups are underway (Figures 4 and 5), but until further reports are available there is hardly any basis for speculation about the reasons for their unique tool-related specialisation.
Development of Tool-oriented Behaviour When observing fluent tool use by an adult New Caledonian crow, it is hard even for seasoned behavioural scientists to avoid interpreting the behaviour in terms of planning and understanding. It is perhaps the presence of these first impressions that highlights the importance of a program of research into the evolutionary, ecological, ontogenetic, and cognitive underpinnings of what the crows do. All classical ethological questions, from causation and development to function and phylogenetic history, are pertinent and hard to answer. Various hypotheses about causation and concomitantly about the reasons why similar behaviour is not observed in other avian species can be used to frame the problem. The following are some possibilities: (a) TOB may result from a set of genetically determined rules, in the sense applied to most animal architecture (e.g., nests, spiders’ webs; Hansell, 1984, 2005); (b) TOB may develop because of particularly advanced reasoning abilities, with individuals ‘working out’ how to solve problems using tools—an inherited capacity for rational insight or a habitat offering special opportunities during ontogeny would explain the rarity of the trait; (c) juveniles may have an inherited tendency to manipulate physical objects at random, gradually converging onto proficient tool use by their own history of reinforcement—in this case, the predisposition for object manipulation (and not for tool use) would be the relevant inherited trait characterising the species; or (d) the behaviour may be passed on through imitation of other tool-using individuals. Hypothesis (d) ignores the problem of how TOB was acquired by the population in the first place and implies that TOB may not be an adaptive specialisation in itself, but a consequence of an enhanced general tendency for social learning. These hypotheses are not mutually exclusive, and it is reasonable to expect that a grain of truth may be associated with each of them and also with other possibilities. One way to eliminate some, however, is to monitor the development of naïve juveniles under controlled conditions. The degree of spontaneous development of behaviour sets boundaries to the need to invoke learning or social transmission and also serves to establish to what extent TOB is an evolved adaptation in this species. Distinguishing between reasoning and a tendency for object manipulation (hypotheses [b] versus [c], respectively) is more difficult, but one might expect a gradual emergence of tool use under hypothesis (c), with non-functional object manipulation persisting after the emergence of successful tool use, whereas reasoning should lead to a sudden acquisition of successful behaviour, with a fast reduction in inefficient behaviours. In the summer of 2004, we reared four crows in our Oxford laboratory (Kenward, Rutz, Weir, & Kacelnik, 2006; Kenward, Weir, Rutz, & Kacelnik, 2005). Three individuals hatched from artificially-incubated eggs, and a fourth was removed from the nest one day after hatching. We investigated the development of two aspects of their TOB: stick tool use, and Pandanus tool manufacture. Development of Tool Use
To make the best use of our limited sample size, we decided to expose different juveniles to different rearing environments. From the time they started to venture outside their artificial nests (a stage called ‘branching’), two of the crows (‘Uek’ and ‘Nalik’) received daily demonstrations of tool use (but not of tool making), in which a human foster parent extracted food from holes and crevices (Figure 6). The other two juveniles (‘Corbeau’ and ‘Oiseau’) never saw tool use, but otherwise received the same degree of overall contact with their human caretakers and, importantly, were fed near holes to control for the possible effect of local enhancement. One of these untutored birds (Corbeau) never saw any tool-like object being handled, but the other subject (Oiseau) may have observed a few accidental instances of a keeper picking-up sticks from the floor of the aviary (although not using them in any purposeful way). By the age of 79 days, all four juveniles retrieved food from crevices with stick tools (Figure 7; Movie 1), with no obvious difference in the onset of tool using between individuals from different treatment groups (Kenward et al., 2005). These observations are sufficient to discard extreme versions of hypotheses (b) and (d) listed above: Although systematic comparisons are still lacking, other corvids such as magpies (Pica pica), western scrub-jays (Aphelocoma californica), European jays (Garrulus glandarius), jackdaws (Corvus monedula), rooks (C. frugilegus), common ravens (C. corax), and American-, carrion-, and jungle crows (C. brachyrhynchos, C. corone, C. macrorhynchos) are often hand-reared for both scientific research or as pets, but none shows a tendency to develop tool use spontaneously. Anecdotal episodes of tool use in wild and captive corvids have been reported (Andersson, 1989; Boswall, 1978, 1983; Caffrey, 2000; Cole, 2004; Gayou, 1982; Jones & Kamil, 1973; Reid, 1982), but all of these are examples of unusual, idiosyncratic behaviour in one or a few individuals (for example, one American crow observed by Caffrey (2000) broke off and used a tapered piece of wood as a tool, of a similar shape to the stepped-cut Pandanus tools made by New Caledonian crows). The time-course and nature of tool-use development was strikingly similar between all of our subjects, including those that were completely untutored. In this regard, the crows appear to be similar to other birds that routinely use tools: Tool use develops spontaneously in woodpecker finches (Cactospiza pallida; Tebbich, Taborsky, Fessl, & Blomqvist, 2001), the only other bird species that habitually uses stick tools in the wild; in Egyptian vultures (Neophron percnopterus; Thouless, Fanshawe, & Bertram, 1989), which use stones to crack open ostrich eggs; and in hyacinth macaws (Anodorhynchus hyacinthinus; Borsari & Ottoni, 2005), which use slivers of wood or leaves as wedges while cracking nuts.
Food retrieval in all hand-reared New Caledonian crows was developmentally preceded by stereotyped object-manipulation patterns (so-called ‘precursor behaviours’) that contained components of the functional, mature behaviour (Kenward et al., 2006). For example, all juveniles engaged in ‘proto-probing’ (first observed in the second week post-branching), where the bird holds a twig in its beak and moves its head back-and-forth in a manner that would be appropriate for probing a hole or crevice to extract food; in this case, however, the twig is moved whilst touching against the surface of another substrate (such as a perch), rather than being inserted into a concavity. Precursor behaviours have also been reported for the development of tool use in the woodpecker finch (Tebbich et al., 2001), and for various other object-oriented behaviours in a range of bird species, including snail-smashing in song thrushes (Turdus philomelos; Henty, 1986), caching in Parids (Clayton, 1992), and nest-building in village weaverbirds (Textor cucullatus; Collias & Collias, 1964). The presence of precursor behaviours in all our hand-reared crows begs the question of whether they play a role in enabling the developing crow to learn about the consequences of object manipulations by honing the functional TOB that develops at later stages. This kind of ‘perception-action’ development is believed to be central to the development of tool-oriented behaviours in the human child and other primates (Gibson & Pick, 2000; Lockman, 2000). The crows’ precursor behaviours did not result in (and hence were not shaped by) reinforcement through food acquisition—all precursor actions were performed weeks before successful food retrieval was observed. This eliminates the possibility that tool use develops as the result of an increased tendency for purely random object manipulation (hypothesis[c]), and emphasises the role of inherited traits together with some form of ‘internal reinforcement.’ Of course, this finding does not reduce the significance of learning, which indeed may itself be strongly based on specific predispositions related to tool use (for further discussion, see Kenward et al., 2006). The development of tool use without social input, the existence of inherited precursor behaviours specific to tool use (namely stereotyped tool-related actions that emerge without reinforcement), and the fact that New Caledonian crows frequently use tools in the wild, indicates that tool use has been specifically selected for and therefore that the behaviour is, or was in the past, an adaptive specialisation. In chimpanzees (Pan troglodytes) and capuchins monkeys (Cebus apella), a long period of object exploration and individual learning precedes functional tool use, with a notable increase in behavioural complexity during development (chimpanzees: Biro et al., 2003; capuchins: Fragaszy & Adams-Curtis, 1997; Lonsdorf, 2005). Furthermore, these species seem to have a predisposition for certain action patterns, such as insertion of objects (capuchins Cebus apella: Fragaszy & Adams-Curtis, 1997; chimpanzees Pan troglodytes: Hayashi & Matsuzawa, 2003). The ontogeny of TOB in crows (Kenward et al., 2006) is similar to that of capuchin monkeys (Fragaszy & Adams-Curtis, 1997) in that non-functional (precursor) behaviours persist until after successful tool use has developed. As a conceptual model, the ontogeny of TOB seems to follow a similar path to the development of song in some passerines and language in humans: Members of the species are endowed with a host of genetic predispositions to acquire the behaviour, but learning—and in the case of song and language, specifically social learning—determines the fate of these predispositions (Marler & Slabbekoorn, 2004). It is thus necessary to consider to what extent social factors can affect the development of TOB in New Caledonian crows. While the onset of tool use did not differ between birds of the two treatment groups, other object-related aspects of development did. Compared to the untutored birds, the two tutored subjects spent more time performing activities related to tool use, such as twig carrying and inserting, but did not differ in measures of general motor development, such as locomotion and carrying of non-food, non-twig items (Kenward et al., 2006). The sensitivity of crows to social factors was further highlighted by the results of a simple choice experiment (Kenward et al., 2006). Tutored juveniles were allowed to observe a human manipulating an object (the object was simply turned in the hand, not associated with any other object) and then given a choice between this and a similar object that had also been present but had not been manipulated by the tutor. Both birds had a strong preference for manipulating the objects that had been handled by the human experimenter. In the wild, such stimulus, local enhancement, or both may play an important role in the acquisition of certain aspects of TOB, such as the choice of appropriate raw materials for tool manufacture. In primates, notably chimpanzees, there is no doubt that social learning does play a significant role in the ontogeny of object-oriented behaviours (e.g., Lonsdorf, 2005, 2006). On the other hand, no effect of tutoring by conspecifics was detected on TOB development in woodpecker finches (Tebbich et al., 2001). Development of Pandanus Tool Manufacture When crows tear a tool out of a Pandanus leaf, a distinctive ‘counterpart’ is left behind (Figures 2b and 3). According to island-wide surveys by Gavin Hunt and colleagues, Pandanus tools conform to three distinct types—narrow, wide, or stepped—and the relative frequencies of these types vary across New Caledonia without any obvious association with relevant environmental factors (Hunt, Corballis, et al., 2006; Hunt & Gray, 2003). This observation constitutes the basis for the suggestion of a cumulative form of cultural transmission of tool technology. Our experimental results show that New Caledonian crows pay attention to the behaviour of others, which is a prerequisite of the culture hypothesis. However, other important pieces of the puzzle are whether socially isolated juvenile crows can successfully manufacture Pandanus tools, and if they do, whether they can produce the different tool shapes observed in the wild. The greater the degree of competence shown by untutored animals, the less we need to invoke social determinants. We first formally tested the response of our hand-reared juveniles to Pandanus leaves when they were between 3 and 4 months old (the two tutored birds were exposed to Pandanus around 2 months old, but were too young to tear at the leaves). To do so, we mounted individual Pandanus leaves on frames in the juveniles’ aviaries, and recorded their behaviour (no birds received demonstrations). All four subjects immediately ripped shreds off the leaves, and one bird (Corbeau) manufactured and used a functional tool on his first day of exposure to the leaves (Movie 2; Kenward et al., 2005). All subjects ripped off leaf fragments that could have been used as tools, although only Corbeau was actually observed extracting food with them. In a subsequent experiment, when the juveniles were between 6 and 7 months old, the two subjects that previously received demonstrations of tool use (Uek and Nalik) were now allowed to observe a human making tools from leaves and using these to retrieve food (but not to keep them or use them themselves). One of the untutored subjects (Corbeau) was given four human-made tools plus intact Pandanus leaves, and the final untutored subject (Oiseau) was exposed to intact leaves but no tools. All subjects received three leaves (mounted more naturalistically than previously, on an artificial ‘tree’) over 5 days, apart from Oiseau, who only received two due to a shortage of available leaves. Functional tools were produced by the tutored subjects (4 tools) and the subject (Corbeau) who had been provided with pre-made tools (12 tools), but not by the subject (Oiseau) simply exposed to intact leaves; all four juveniles continued to produce non-functional strips (Kenward, 2006). This is consistent with the result of the first experiment: namely, that captive-born New Caledonian crows will tear tool-shaped strips from Pandanus leaves in the absence of social inputs, and hence without a requirement for culture. However, the pieces removed from the leaves and used as tools were crude in shape and manufactured using a different technique from that used by (at least) one wild crow observed by Hunt and Gray (2004b). Therefore, the possibility remains that social influence is important in promoting and refining Pandanus tool manufacture in wild crows. The failure to produce refined stepped tools could have occurred for several reasons, some of which are supportive of the culture hypothesis while others argue against it. Reasons why invoking culture may not be necessary include the following:
In support of the culture hypothesis, it could be that, while some aspects of TOB are narrowly canalised genetically, social learning is a prerequisite for the acquisition of specific manufacturing techniques, tool designs, or both. These options are clearly separable with further data, and forthcoming research will clarify the issue. Social learning need not involve faithful imitation of the exact motor actions exhibited by tool-producing conspecifics. More rudimentary social processes such as stimulus or local enhancement (Galef, 1988; Heyes, 1994), as demonstrated in our choice experiment mentioned above, could expose wild juveniles to a biased set of raw materials and hidden prey. Juveniles may also learn about particular Pandanus tool shapes from inspecting abandoned tools or tool counterparts in leaves, without ever observing another bird manufacturing a tool. In other words, the hypothesis of a tool culture is compatible with various social transmission mechanisms, ranging from elaborate cognitive properties to better-known and widespread learning processes. It is interesting to view our experimental findings in the light of first results on juvenile development in the wild. We have monitored radio-tagged wild crows from 2005 onwards, and our preliminary data suggest that juveniles stay in (extended) family groups for at least the first year of life, foraging often in close association with parents, siblings, and other individuals. This period of dependency prior to natal dispersal could provide opportunities for social transmission of information: vertically (from parents to offspring), obliquely (from the parental to other members of the offspring generation), or horizontally (between juveniles of different ages). There are also very suggestive observations of juveniles watching their parents using tools and subsequently using the same tools (Hunt, 2000b; Sarsby, 1998). In conclusion, proficient TOB in New Caledonian crows results from a complex interplay of heritable predispositions, individual learning through object exploration, and (quite possibly) the acquisition of socially-transmitted information. In conjunction with further work on captive subjects, the long-term observation of individually identifiable wild subjects will be instrumental for teasing apart the relative contributions of these different mechanisms. Deployment of Tool-oriented Behaviour The fact that at least some aspects of New Caledonian crows’ tool-oriented behaviour develop from unlearned, heritable predispositions does not imply that TOB must be stereotyped or unrelated to reasoning in adult crows. Consider again our earlier analogy with the development of human language. A prevalent view argues that there is an inherited basis for the mechanisms of human language acquisition (Vargha-Khadem, Gadian, Copp, & Mishkin, 2005; but see Fitch, Hauser, & Chomsky, 2005), but it would be unreasonable to maintain that cognition is absent from conversation. Tools are an unusual way of interacting with the physical world. Therefore, independent investigation is necessary to explore the cognitive underpinnings of TOB deployment by adult crows—their folk physics, or "common sense understanding of how the world works and why it works in the way it does" (Povinelli, 2000, p. 9). It is worth noting that, while tool use may be favoured by the pre-existence of a high level of understanding (as some argue for humans; e.g., Wolpert, 2003), the same behaviour, whatever its control mechanism, may promote the evolutionary development of folk physics by exposing the subjects to situations where understanding the causal basis of interactions between objects gives them a competitive advantage, thus reversing the arrow of causality. Another possibility is that exposure to extensive object-object interactions during a juvenile period promotes the development of folk physics and that this tendency to play with objects is the species-specific cognitive adaptation responsible for the unique level of tool use. In the next section, we first examine how selective and flexible adult crows are in making and using tools, providing hitherto unpublished re-analyses of data, and then we consider whether crows may possess anything worthy of the term ‘understanding.’ Choosing Tools A fundamental feature of intelligence is flexibility (Emery & Clayton, 2004b; Lefebvre, Reader, & Sol, 2004; Reader, Sol, & Lefebvre, 2005; Roth & Dicke, 2005): being able to alter one’s behaviour if circumstances change (John Maynard Keynes famously replied to a fellow parliamentarian who accused him of inconsistency that "When the facts change, I change my mind. What do you do, sir?"). Particularly diagnostic of intelligence is the ability to alter one’s behaviour adaptively: that is, to assess how to change one’s behaviour to cope best with the change in circumstances. Therefore, perhaps the first question to ask of a tool user is whether s/he can choose the best tool out of a selection of tools of different dimensions when the task is varied. If tool use is based on rigid rules for choosing particular tools in particular situations, then we would expect that, when confronted with novel problems (i.e., those not typically encountered in natural contexts), subjects would continue with their default behaviour or choose at random. Choice of the correct tools in novel tasks would at least indicate that the natural behaviour was not rigidly specified. It is less clear whether such flexibility indicates the involvement of reasoning or abstract concepts, although, depending on the task, choice could be governed by previously learned associations between particular tools and success or by an understanding of the relationship between tool dimensions and task demands. Wild chimpanzees use tools of different dimensions for different tasks (e.g., Boesch & Boesch, 1990, 1993; Hicks, Fouts, & Fouts, 2005; Nishida, 1972). This has been taken as evidence for selectivity and ‘mental representation’ of task demands (Boesch & Boesch, 1993; Visalberghi, 1993). However, since the chimpanzees were not described as solving novel problems, it is difficult to assess the extent of individual flexibility. In controlled experiments, captive gorillas (Gorilla gorilla) and orangutans (Pongo pygmaeus) appropriately chose the longer of two tools to rake in a food reward when it was distant and chose randomly between the tools when the reward was close (Mulcahy, Call, & Dunbar, 2005; see also Pouydebat, Berge, Gorce, & Coppens, 2005), as did a Tonkean macaque (Macaca tonkeana) in a similar task (Ueno & Fujita, 1998). Two capuchins also spontaneously chose the thinner of two sticks when necessary (Anderson & Henneman, 1994). In contrast, woodpecker finches did not spontaneously choose longer tools when food was further away (Tebbich & Bshary, 2004). In Tebbich and Bshary’s experiments, five birds were given sticks of five different lengths and food at five different distances inside a horizontal transparent tube. On their first trials, all subjects chose short tools, and across all trials the length of the tools they first used did not correlate with the distance to food, although over the course of the experiment three subjects did appear to choose sufficiently long tools more frequently than expected by chance (Tebbich & Bshary, 2004).
A relevant question is, therefore, whether New Caledonian crows can choose tools of appropriate dimensions for novel problems. This was first assessed by giving two subjects (‘Betty’ and ‘Abel’) tools of ten different lengths and food at different depths inside a transparent, horizontal tube (Chappell & Kacelnik, 2002). Both subjects selected appropriate tools from their first trials and chose tools that were too short to reach food on just 13% of trials, against a chance expectation of 45%. Each subject chose the tool that exactly matched the distance to food on 5 of their first 20 trials and the longest available tool on 10 trials (both significantly greater than expected by chance). In addition, new analysis of the data reveals that, excluding all trials in which the longest tool was chosen, there is a significant increase in minimum tool length chosen by each subject at each distance to food (Figure 8; Spearman rank correlation; for Betty [who participated in only one experiment], N = 7, rS = 0.764, p < 0.05; for Abel [who participated in two experiments], on pooled results from both experiments N = 10, rS = 0.652, p < 0.05]). The subjects were also tested on a task where they had to select tools of the appropriate width. Here, a tool could be inserted through a hole of three possible diameters to dislodge a food reward (Chappell & Kacelnik, 2004). In the first experiment, Betty was given three tools of different diameters (1 mm less than the diameter of the three holes) in different configurations: either all loosely tied into one bundle, or two tied together and one free. Betty showed a strong preference for the tool with the smallest diameter, to the point of dismantling the bundle to retrieve it even when a suitable tool was freely available. That she would incur this additional handling cost suggests selectivity (she strongly preferred one tool) at the expense of flexibility (other tools were suitable, but she did not use them). In a control condition, where all the tools were available and suitable for food-retrieval, Betty showed the same preference, suggesting that her affinity for the narrowest tool may have been driven by ergonomic factors.
The previous test was concerned with tool use, but we also conducted experiments to investigate aspects of our crows’ tool manufacture. Both Betty and Abel were tested together on the same apparatus as used for the width-selectivity test, but with access to raw materials for tool manufacture (an oak branch; Movie 3) rather than artificial tools. In each trial, the subjects approached the tube and looked at it before making tools. Out of 30 trials, the male (Abel) retrieved food on 13 and the female (Betty) on 16 (the remaining trial was aborted because neither subject had obtained the food after 30 minutes). Abel retrieved food using the first tool he made (without modifying it) in 13 trials, only making an inappropriate tool on 1 trial (the tool was narrow enough, but too short to dislodge the food). Betty retrieved food using the first (unmodified) tool she made on 10 trials, and on 2 trials she modified the tool to make it narrower after trying to use it (Movie 4). On 5 trials, her first tool was too short, although suitably narrow, and on 4 of these she made another tool that she used to retrieve food. Pooling results from both subjects, the maximum diameter of the (unmodified) tools increased significantly with the diameter of the hole (Figure 9a; Chappell & Kacelnik, 2004). Strikingly, 16 of the 24 tools made in middle- and large-hole trials would have been too thick if used on small-hole trials, whereas only one tool made in the narrow-hole trials was too thick (and it was subsequently modified to make it narrower). There is no quantitative information on how the crows may benefit from selectivity in this task (as opposed to making exclusively narrow tools that are functional for all three hole diameters), but a post hoc analysis of one aspect of the results not examined in the original publication reveals a potential and perhaps non-obvious association between tool manufacture and foraging efficiency. Whilst the latency to success (from the start of the trial) showed no trend with hole diameter, this latency decreased with diameter of the manufactured tool (only including trials where the first tool made was successfully used, unmodified; Figure 9b; GLM analysis with ‘latency to success’ [transformed by reciprocal square root] as the dependent variable, tool diameter as a continuous explanatory variable, and subject as a random explanatory variable; for tool diameter, F1,18 = 7.95, p = 0.011; for subject, F1,18 = 0.04, p = 0.848, F1,24 = 0.18, p = 0.671]. This is consistent with the hypothesis that the subjects were being selective only when required since the narrower the desired tool diameter, the fewer the tools that fulfil the criterion and the longer the requisite search time until a suitable twig is found. Thus, crows seem capable of manufacturing appropriately-sized tools from raw materials as well as of choosing from a set of available tools according to the present needs, although their choice may also be constrained by inherent preferences, whether these are the product of previous learning or simply ergonomic limits. A somewhat different picture of flexibility emerges from field experiments. Hunt, Rutledge, and Gray (2006) examined how two free-living New Caledonian crows made (or chose) and used tools to extract food from an artificial container—either a vertical box with a transparent side, or opaque vertical holes in a tree stump. Food was placed at one of two depths, which varied between blocks of trials. One subject was given Cerambycidae larvae (Agrianome fairmairei, provided alive or dead; see Figure 5, top panel) and used mainly leaf stems to extract them, while the other manufactured Pandanus tools to extract supplied lumps of meat. The main finding was that the depth of the hole did not affect the length of the first tool with which either crow attempted to extract food (this tool tended to be shorter than the distance to food in the deeper holes), and only after failure to retrieve food with the first tools did the subjects make or select longer tools. Therefore, there was no evidence that the subjects assessed the distance to the food before choosing tools, although they did adapt their behaviour once the tools had been tried. The authors argue that this demonstrated that the crows "may generally use a two-stage heuristic strategy to solve tool problems" (Hunt, Rutledge, et al., 2006, p.307)—namely, initially using default behaviour and modifying this appropriately following failure. They argue that this rules out the possibility that the crows were using ‘immediate causal inference’ to solve the problem and that the adaptive modification of tool length following failure was consistent with either a ‘previously developed associative learning rule’ or ‘delayed causal inference’ (Hunt, Rutledge, et al., 2006). These observations contrast with those of Chappell and Kacelnik (2002, 2004) described above, who found selectivity and flexibility for length and diameter from first attempts in a new task. In our opinion, however, major methodological differences between the studies preclude direct comparisons. Of particular relevance is the fact that the study by Hunt, Rutledge, et al. (2006) used free-living crows and a naturalistic foraging task. Consequently, experimental trials were embedded within a natural foraging context, where the crows (presumably) routinely used tools. Free-living birds are likely to develop a preference for using a tool length that is sufficient to reach prey in most natural holes, a view that is supported by our own ongoing work on stick-tool use in wild crows. If this is the case, crows may have approached the task in Hunt, Rutledge, et al.’s experiment with their default behaviour, not noticing that it required a different kind of tool (the ‘two-stage heuristic strategy,’ in Hunt, Rutledge, et al.’s terminology). In contrast, in Chappell and Kacelnik’s laboratory studies, experimental trials represented the majority of the daily tool-related foraging by the subjects, and the novel nature of the task (the distance to food varied between consecutive trials, rather than between blocks of trials, and food was presented in a highly distinct, novel apparatus) may have elicited case-by-case decision making. The nature of the apparatus (vertical opaque- or partially-opaque-sided holes for Hunt, Rutledge, et al.’s subjects; a horizontal transparent tube for the Oxford subjects) also provides a possible explanation for the different results. The subjects in Hunt, Rutledge, et al.’s experiments may not have exposed their ability to discriminate before choosing the initial tools because they simply were not able to assess the distance to food in vertical, opaque, or semi-opaque holes. Other methodological differences hamper comparisons. For instance, Hunt, Rutledge, et al.’s study used two food depths in separate blocks of trials, whereas Chappell and Kacelnik’s (2002) experiment exposed subjects to 10 distances to food in randomly interspersed trials. These disparities highlight the complementarity of field and laboratory studies. While the former are irreplaceable for investigating ecological relevance, the latter allow for stricter control of the subjects’ state and experience and demonstrate abilities that may or may not be important under natural circumstances. No doubt, both approaches are necessary to elucidate detailed mechanisms of learning and choice. In conclusion, New Caledonian crows, like some tool-using primates, are able to select appropriate tools for novel tasks. In the laboratory, they make and use tools from a variety of materials, including (their own moulted) feathers, cardboard, and wood chips, and they readily use any elongated object for probing holes and crevices, e.g., screws, strips of plastic, or wire (Kacelnik, Chappell, Weir, & Kenward, 2006; personal observations). Appropriate selection of tools based on length has not as yet been demonstrated for wild crows (Hunt, Rutledge, et al., 2006), or in a similar experiment in captivity with the other avian stick-tool user—the woodpecker finch (Tebbich & Bshary, 2004). While selectivity and flexibility are to be expected if behaviour is controlled by logical inference and planning, their presence does not prove causal understanding. For example, subjects could have a rich repertoire of previous associations between tools of particular dimensions and holes of a certain depth or diameter, which would enable them to choose correctly in ‘novel’ tasks. Selectivity—defined as the use or manufacturing of tools conditional in size or shape on the task being faced—raises the question of which cognitive mechanisms are employed in TOB, but it is not, per se, sufficient to identify any particular ability that excludes conventional processes such as associative learning plus generalisation. Understanding? As mentioned in the introduction, humans possess intuitive abstract ‘theories’ about the way the physical world operates, incorporating a variety of principles (such as gravity, force, connectivity, weight) that allow us to operate as obligate tool users in a materially complex environment and to predict how objects will interact in novel situations. While this can be described as causal understanding of the physical world, it need not equate to formal understanding in any rigorous scientific or philosophical sense. For example, by 12 years of age, children (who presumably have experience using levers in their everyday life) are very proficient when asked to judge how the position of weights along a scale will affect the balance of a lever (for many, a novel task) (Amsel, Goodman, Savoie, & Clark, 1996), yet they lack a formal understanding of the underlying principles (that the mass multiplied by the distance must be equal on both sides of the fulcrum) until this is explicitly taught (Stephen Barlow, personal communication). Likewise, through experience most people have an implicit knowledge of what makes an effective hammer. If one forgets to bring a hammer on a camping trip, one can choose another suitable object with which to knock in the tent pegs regardless of superficial visual dissimilarities. This same rationale—a task requiring transfer of concepts derived from past experience to a novel problem—can be used to attempt to assess causal understanding in non-humans. To test for causal understanding, two main experimental approaches have been used to date. The first is another kind of selectivity test: Subjects are offered choices either between different tools (where effective tools share a common mechanism—such as ‘hooking’—but are perceptually different), or between different actions with a tool (such as pushing or pulling a tool, or choosing which side of an apparatus to insert a tool into). Subjects are usually trained on a particular configuration of the apparatus and are then tested with the task or tools transformed in a manner which alters the causality of the problem. Transformations are typically designed such that subjects who have learned ‘procedural rules’ (sensu Povinelli, 2000) based upon superficial characteristics of the apparatus (such as colour, shape, texture) will behave in one way, whereas those that have formed a concept about how the apparatus functions (for example, that the tool needs to be connected to the food in order to act on it) will behave in another way. However, since the number of options the subjects face is normally limited, there is a relatively high chance that they will perform correctly by chance. Therefore, a robust study requires either a large sample of subjects to be tested, or a suite of different tasks or transformations to be used. Furthermore, the limited number of options and trials make one-trial learning very plausible, so relatively little can be inferred from post-first-trial performance. For example, in a task with only two options a win-stay lose-shift strategy would lead to perfect performance after the first trial. The second approach focuses on the animals’ spontaneous behaviour. Rather than giving subjects a defined set of choices, they are placed in a situation where they have a low probability of solving a task by chance alone (for example, in a hook making task an animal may be given a piece of pliable material that can be changed into an infinite number of shapes, but only a small subset of these shapes would be functional). Subjects with no causal understanding might be expected to depend on trial-and-error and, therefore, to follow a gradual pattern of acquisition similar to that in classical operant shaping experiments (e.g., Thorndike, 1898). The absence of insight should lead to gradual performance improvement because success is achieved following a sequence of behaviours, any one of which (from the subject’s perspective) could be the critical element. In contrast, subjects that acquire and apply general principles might equally rely on random exploration until success but then infer the reasons for success and organise their future behaviour towards this goal, hence improving in a more step-wise pattern. Experiments with richer tasks, however, suffer from difficulties in formulating null hypotheses and their respective probabilities (for example, how can one define the likelihood of manufacturing a hook-shaped instrument to extract a reward by chance alone?), and in comparing performances across species. Animals differ in their behavioural repertoires and, therefore, in their likelihood of solving given experimental tasks (cows, for example, seem unlikely candidates for bending wire into hooks, despite reports to the contrary by a British newspaper; Townsend, 2005). Both approaches, therefore, have their advantages and should be seen as complementary, but the limitations of each of them should be taken into account when interpreting results. We will return to the issue of what conclusions can be drawn from such work in Section 6. We now turn our attention to the performance of New Caledonian crows and other animals in the well known ‘trap-tube’ test and discuss what can be deduced from these results regarding the presence of physical understanding. The Trap-tube Test The most widely applied test for causal understanding in the physical domain is the trap-tube task (Visalberghi & Limongelli, 1994). In its simplest form, it consists of a transparent horizontal tube with a recess (‘trap’) in the middle. Food is placed in the tube and subjects are given a stick with which to retrieve it. In the ‘active’ state the trap is oriented downwards; if the subject moves the food across the central part of the tube, the food falls into the hole and becomes trapped. In the ‘inactive’ state, the trap is oriented upwards; here the subject can move the food either way with impunity. Logically, a subject who understands gravity should never move the food across the central part of an active tube, but should show no such bias when faced with an inactive tube. In some experiments another transfer test has been used, where post-training the trap is positioned off-centre to test for use of a distance-based associative rule (e.g., Limongelli, Boysen, & Visalberghi, 1995). The only New Caledonian crow tested to date (Betty) did not instantly solve the task, but instead she improved gradually. After around 60 trials, she spontaneously developed a two-stage technique. This involved first inserting the stick in the tube from the ‘safe’ side (leaving the trap between herself and the food) until its distal end protruded from the opposite side, then walking to the opposite side and pulling the stick so as to extract the food in a controlled manner. This technique avoided the trap and had the added benefit of eliminating the danger of another crow snatching the food, an event that could occur when pushing the food out of the opposite side of the tube. Using a combination of this technique and a standard pulling action (pulling the food towards her from the side closest to the food), she reached criterion (obtaining the food on 8 or more trials in three consecutive sessions of 10 trials) after 110 trials (Movie 5; Chappell & Kacelnik, 2007; Kacelnik et al., 2006). This time to acquisition is comparable to that of chimpanzees (Limongelli et al., 1995; Reaux & Povinelli, 2000), capuchins (Visalberghi & Limongelli, 1994) and woodpecker finches (Tebbich & Bshary, 2004) tested on the same task, but comparability is difficult because of Betty’s use of the two-stage technique. When the trap was switched to the inactive state, Betty continued to avoid it using her technique, even though it was now irrelevant. Chimpanzees (Reaux & Povinelli, 2000) and capuchins (Visalberghi & Limongelli, 1994) also continue to avoid the inactive trap, and consequently it has been argued that they do not understand the causal basis of the task. In contrast, one woodpecker finch reverted to chance behaviour in the transfer test (Tebbich, Taborsky, Fessl, Dvorak, & Winkler, 2004). Two chimpanzees and a capuchin monkey have been tested with the off-centre trap design, following successful performance with the centred trap: The capuchin now performed significantly below chance, demonstrating that she was probably using a distance-based associative rule, whereas the two chimpanzees remained highly successful (Limongelli et al., 1995). Interesting as it is to compare performance levels across species, the conclusions that can be drawn from this general experimental design are limited (Chappell, 2006). One caveat is that it is not clear what the cognitive implications are of failure to revert to random insertion after the trap is inverted (Machado & Silva, 2003). Although logically it is not necessary for the subject to carry on avoiding the ineffective trap, if there is no cost to doing so (as in most published experiments) even subjects who are capable of understanding might find no incentive to modify their behaviour. Interestingly, even humans continue to avoid the inverted trap, despite their understanding of the physical principles involved (Silva, Page, & Silva, 2005). It is equally hard to draw conclusions from observations where subjects do revert to random tool insertion with the inverted trap. It is possible that such subjects had learned through reinforcement the characteristics of the stimulus (i.e., the tube with downward-oriented trap) so specifically that they fail to generalise when the trap is inverted, and they react as if in a completely new task, to which they therefore respond randomly. Alternatively, subjects might monitor the moment-to-moment movement of the food with respect to the trap; if the trap is inverted, the food will never approach it closely, and subjects will therefore insert tools randomly. This is thought to account for the performance of the one woodpecker finch that ‘passed’ the transfer test (Tebbich et al., 2004). There are similar problems with the off-centre trap test. Although below-chance performance indicates that subjects were using a distance-based associative rule, above-chance performance does not mean that they understood the causal basis of the task. They might have been using another associatively-learned rule, such as ‘push the food away from the trap,’ without any understanding as to why they should do this. To solve problems associated with the standard task, Nicky Clayton and colleagues (Seed, Tebbich, Emery, & Clayton, 2006; Tebbich, Seed, Emery, & Clayton, in press) developed an alternative task to test the same principle, but which could also be used by non-tool-using species—in their experiments, rooks. To adapt the task to non-tool-users, wooden dowels with two solid transparent disks attached (with the food reward positioned between them) were pre-inserted into tubes, so subjects simply had to pull the dowel to retrieve the reward. Various configurations of tubes were then used to test the subjects. In the study by Tebbich et al. (in press), following training to pull the dowel out of a plain, horizontal tube to get food, eight subjects were tested with a tube with a single, off-centre trap. Those subjects that reached the criterion for success on this task were then tested with a tube where food was placed between two apparent ‘traps,’ only one of which was functional at any time. This tube had two possible configurations (see Figure 10): In one (Tube A), a black disc blocked one trap at the bottom and the other at the top, so the subject had to move the food over the latter trap and out the mouth of the tube; in the other (Tube B), one trap was again blocked at the bottom by a black disc, but the second trap was open, so food could be retrieved by pulling the dowel towards the trap with no base, upon which the food would fall clear of the apparatus. In both tube types, therefore, the active trap featured a black disc at its base. Half the subjects were tested with Tube A first, and half with Tube B, receiving between 40 and 50 trials with each. Three of the subjects reached criterion performance (after 30, 40, and 50 trials) on the single-trap tube, but all remained at chance throughout testing with both two-trap tubes.
Seed et al. (2006) tested a separate group of eight rooks in a very similar manner. Crucially however, they did not first train the subjects on the single-trap tube. Instead, half the subjects were trained with Tube A, and half with Tube B. The seven subjects that reached criterion were tested on the alternative configuration (i.e., those that received Tube A were tested on Tube B, and vice versa). All 7 rooks were successful in extracting food in their first trial with the new configuration, and they were successful more often than expected by chance over 20 trials. These results are consistent with the subjects learning the causal features of the task but could also be explained by learning to avoid traps with black discs at the bottom. To exclude the latter possibility, further transfer tests were administered (see Figure 10). In Tube C, one trap was blocked at the top and the other one had no base, and both ends of the tube were blocked by bungs; food could be extracted through the baseless trap, but subjects could only pull the stick once, as after the first pull the distal end of the stick disappeared into the bung and could not be pulled out again. Tube D had the same traps but no bungs in the ends of the tube; however, the apparatus was lowered to the ground such that food could only be extracted by pulling over the trap with the blocked top. These tubes were designed so that the stimuli that previously signalled which direction to pull the stick (such as "pull away from a black disc at the bottom of trap") were changed or removed, so that subjects responding on the basis of a single associatively-learned cue would perform at or below chance. Subjects were exposed to two blocks of ten trials of Tube C, followed by two blocks of ten trials for Tube D, or vice versa. Six subjects were at chance on Tubes C and D, but one rook, ‘Guillem’, showed near-perfect performance on both from his first trial. While appropriately cautious of conclusions based on a single subject, Seed et al. (2006) propose that Guillem possessed either an understanding of the observable forces involved, or had abstracted the rules of the task (see also Chappell, 2006). However, success on any one task, particularly one involving a choice between just two defined actions, could be because the subject happened, by chance, to learn about cues that were causally-relevant for that task. For example, Guillem could have learned that two cues predicted success: When the mouth of the tube is open, pull the food towards the trap with the black disc on the top; and pull the food towards a trap that has no black disc, if there is space underneath the tube. The striking individual differences in performance (only 3 of 7 rooks learned to solve the original trap-tube task in Tebbich et al., in press, and none passed the transfer tasks; Guillem was the only one of 8 subjects to pass the transfer tasks in Seed et al., 2006) make this possibility more plausible, since if rooks were predisposed to form theories about physical causality, we would expect more of them to demonstrate this ability in the experiments. It is worth comparing this work with that on capuchin monkeys. The performance of capuchin monkeys seems to be based almost entirely on trial-and-error learning of procedural rules (Visalberghi & Limongelli, 1994; Visalberghi & Trinca, 1989). In some experiments, chimpanzees have shown greater sensitivity to the causally-relevant aspects of the tasks (e.g., Limongelli et al., 1995; Visalberghi, Fragaszy, & Savage-Rumbaugh, 1995), but these successful performances fall short of demonstrating an understanding of the physical mechanisms involved (see Povinelli, 2000). Sensitivity to functionally-relevant features of tools has also been demonstrated by a series of experiments by Marc Hauser and colleagues, primarily with non-tool-using cotton-top tamarins (Saguinus oedipus; Hauser, 1997; Hauser, Kralik, & Botto-Mahan, 1999; Hauser, Pearson, & Seelig, 2002; Hauser, Santos, Spaepen, & Pearson, 2002; Santos, Rosati, Sproul, Spaulding, & Hauser, 2005), but also by testing rhesus macaques (Macaca mulatta; Santos, Miller, & Hauser, 2003), common marmosets (Callithrix jacchus; Spaulding & Hauser, 2005), and vervet monkeys (Cercopithecus aethiops; Santos, Pearson, Spaepen, Tsao, & Hauser, 2006). However, these results suggest that the subjects possess domain-specific biases regarding what features of objects are relevant in certain situations, rather than an understanding of underlying causality (Penn & Povinelli, 2007). Indeed, in recent experiments Santos and colleagues have tested tamarins on experiments similar to those used by Povinelli and colleagues with chimpanzees (Povinelli, 2000) and apparently found that like chimpanzees, the tamarins failed to distinguish between the functional and non-functional tools or actions (reviewed by Hauser & Santos, in press). In conclusion, the result from these experiments, and others employing a similar strategy of offering subjects choices between tools or discrete actions (e.g., Povinelli, 2000, chap. 4-10; Visalberghi & Trinca, 1989), have yielded, at best, very limited evidence for understanding of physical causality in non-humans. Hook Making As discussed above, another technique for investigating non-humans’ understanding of physical causality is to observe their problem-solving behaviour in situations that are too rich to be fully defined by a choice. To this end, we have examined how New Caledonian crows manipulate pliant material to form functional tools.
This line originates from an unplanned observation in an experiment where Betty and another crow, Abel, faced a choice between a straight or a hooked piece of garden wire, with food available in a small bucket at the bottom of a vertical transparent tube (Weir, 2006). The goal was to extend the tests of flexibility and selectivity (see Weir, 2006, Section 4.1) to include tool shape in addition to length and diameter. The apparatus was designed such that the hooked wire was functional for retrieval of the bucket, but the straight wire was not (this last restriction proved to be almost, but not absolutely effective). On the fifth trial of this experiment, Abel probed into the tube with the hooked wire but then flew off with it to another part of the aviary before retrieving the bucket. Betty attempted to lift the bucket with the straight wire, and when this proved ineffective, proceeded to bend it into a hook, which she used to extract the bucket (Figure 11; Weir, Chappell, & Kacelnik, 2002). Although this has become a textbook example of animal intelligence (e.g., Barnard, 2004; Boyd & Silk, 2006; Freeman, 2002) and was termed "the most impressive example of folk physics in any animal" in a recent review of bird cognition (Emery, 2006, p. 32; note that Emery wrote this before the publication of the results of Seed et al., 2006), unravelling the cognitive processes behind it presents serious challenges. Perhaps the crucial issue is whether Betty’s wire-bending demonstrates causal understanding. In 10 trials following the initial observation, Betty was only given a straight wire and nearly always briefly attempted to retrieve the bucket with the unmodified tool (Movie 6), but then she consistently bent the distal end of it (i.e., the end not held in her beak) using two different techniques both involving fixing the tip of the wire and pulling laterally from the proximal end (Weir et al., 2002). The bent part at the distal end formed the hook. (Note that we are using the term ‘hook’ informally to describe a bent piece of metal that could function to pull the bucket out of the well; we recognise that, as pointed out by Emery, 2006, in several cases the angle of the bend was less than 90 degrees.) Betty had been wild-caught as an immature bird two years before the experiment took place, so her experience before entering captivity is unknown, but we are not aware of any natural materials that could be bent and used like garden wire. To our knowledge, Betty had never experienced wire or similarly pliant objects whilst in captivity, apart from 1 hour of free manipulation with flexible pipe-cleaners a year before this experiment (which she was not seen to bend or use as tools). In subsequent experiments, Betty was presented with the same apparatus but a new material for tool making: flat strips of aluminium. On most trials, she first attempted to retrieve the bucket with the unmodified material, but the duration of these attempted probes declined rapidly (the median duration was just 3 seconds). After these attempts with the straight strips, in the first few trials she attempted to make distal bends, following her previous techniques (Weir & Kacelnik, 2006), but because of the properties of the new material she was unsuccessful. (The strips could not be wedged in a substrate in the same manner as the wire.) Betty then developed a new technique, proximal bending, that was more effective with the aluminium and which she used on all subsequent occasions (Figure 12a; Movie 7). This involved bending the end of the strip that was held in her beak, rather than the distal end as previously. A consequence of this technique was that after modification she was holding the modified end of the tool, so the instrument needed to be turned around before it could be used. However, in 5 of the first 10 valid trials, she initially attempted to retrieve the bucket with the unmodified end of the strip whilst holding the hooked end.
Interpreting the significance of Betty’s behaviour in terms of causal understanding is not straightforward. One approach is to ask what behaviour we would expect from an agent capable solely of trial-and-error learning. Such an agent would presumably have learned in the first experiment (Weir et al., 2002) that a certain sequence of actions with the wire leads to success: First, the distal end of the wire must be wedged, then the proximal end pulled at an angle at the distal end, then the wire removed and inserted into the tube and used to pull up the bucket. Since the new task and material closely resembled the old one, we would expect such an agent to perform initially the same series of actions—which is indeed what we observed with Betty. Following failure of this sequence, the agent’s behaviour should become more variable (since variability increases in extinction: e.g., Neuringer, Kornell, & Olufs, 2001), but in a (relatively) random fashion. This was not what happened with Betty: Her behaviour did become more variable, but it focussed specifically upon manipulating the shape of the tool, rather than on other components of the previously-successful sequence, such as the tool insertion or lifting action. Therefore, it seems that Betty’s bending actions were to some extent goal directed in that she was intent on producing a hook or hook-like modification. It also appears that she had some understanding of how to make a hook, since she very quickly converged on an efficient, novel way of doing so. In other words, in both experiments she was able to invent a method of shaping the material into a functional and probably preconceived form. However, these results suffer from the problem raised above: namely, that it is inherently difficult to pose a null hypothesis against which to test these results. We are, therefore, unable to quantify the statistical significance of our observations, highlighting a fundamental problem in the investigation of creative behaviour. Betty’s repeated attempts to use the wrong end of the modified strips suggest that she did not understand exactly why a hook was needed— certainly, an agent who fully understood the task should never probe with the wrong end of the tool. However, we should be cautious about leaping to judgements of intellectual inability based on inefficient performance since humans do sometimes make similar mistakes despite undoubtedly possessing sophisticated folk physics (discussed in more detail later in this section). It is also interesting to note that chimpanzees only correctly reoriented a hooked tool on 6 of 28 trials (4 trials per subject, 7 subjects), which was the same as the number of times they reoriented a straight tool (Povinelli, Reaux, Theall, & Giambrone, 2000b, Experiment 16 conditions E and G). We tested Betty’s understanding of the relationship between tool-shape and success with two additional brief experiments involving more radical transformations of the tasks (Weir & Kacelnik, 2006). In the first, the aluminium strip was pre-bent at both ends, preventing it from being inserted through a narrow hole—an action which was necessary to dislodge food. Betty did modify the tool and obtain food on the only valid trial (in two trials she managed to retrieve food without modifying the tool), although we cannot conclude that this action was goal-directed. She flattened both ends of the tool (Figure 12b), which may have been for ergonomic reasons (ease of holding) rather than a purposeful modification of the ends to serve two distinct functions (proximal end to be held in the beak, distal one to fit the hole). In the second experiment, the aluminium strip was provided pre-bent into a broad U-shape, which was too short for the task; it had to be lengthened by unbending to retrieve food from a horizontal tube. Betty unbent the tool and obtained the food on two of the three valid trials (again, Betty retrieved food on one trial in a non-anticipated manner; Figure 12c; Movie 8), although on both occasions she first attempted to reach the food with the unmodified tool. Sadly, Betty died before completing more replicates, and the small number of trials together with the variability in her behaviour (to be expected in this kind of experiment) precludes statistical analysis. However, it is important to note that she never performed a bending action in these tasks, even though this had been the behaviour consistently deployed and rewarded in previous experiments. Thus, while her ability to ‘design by reasoning’ cannot be asserted, it is reasonable to doubt a simple associative-learning account: In spite of her reinforcement history, bending was not her generalised response to inaccessible food and manipulable material (Figure 12). Although Betty did not show a human-level understanding of the causal need for a hook, her degree of conceptualisation of the problem seems to exceed that previously documented in other animals. Of particular relevance are a series of tool-modification experiments with captive chimpanzees (Povinelli, Reaux, Theall, & Giambrone, 2000a). In these studies, 7 chimpanzees were each given 12 trials where they had to unbend a tool to retrieve the reward, interspersed with 18 probe trials where no modification was necessary. Only one subject (‘Jadine’) ever successfully unbent the tool when it was necessary to do so, and she did so only on the final four test trials following explicit training (to bend the tool). Moreover, when only one end of the tool was bent, many of the subjects preferentially attempted to insert the wrong end into the apparatus throughout the testing period. These results suggest that chimpanzees can learn (in this case through explicit provision of information) the properties of tools while remaining ignorant of the abstract concepts involved in successful tool use.
What the experiments with Betty do confirm is that understanding does not conform to a presence/absence dichotomy. Some anecdotal observations with humans may serve to illustrate this point. We recently presented our research on New Caledonian crows at the Royal Society of London’s ‘Summer Science Exhibition.’ We asked some visitors to our stall—adults and teenagers, including many Fellows of the Royal Society and even its most recent past President (Figure 13)—to participate in an informal experiment resembling that conducted with Betty and Abel. Subjects were given the opportunity to extract souvenir badges from a vertical, transparent tube, using a pair of pliers (to simulate a crow’s beak) and a piece of straight wire or aluminium strip. We explained that the goal of the task was to extract the badge, and most visitors had seen video footage of Betty’s successful wire bending. Surprisingly, and like Betty had done during early trials, a substantial proportion of people attempted to retrieve the reward without modifying the starting material—an approach that was only successful in a small fraction of attempts. Many other participants produced an exaggerated hook with a U-shaped tip that was unsuitable for the desired task. In a few particularly revealing cases, subjects bent a suitable hook—probably imitating Betty—but then inserted the straight, non-functional end into the tube! This exercise had a pedagogical and slightly humorous intention, and is hardly a rigorous test of humans’ ability on this task. Yet, despite their definite understanding of the goal as well as their likely knowledge of the pliability of wire, many humans did not deduce the utility of a hook or design it correctly before engaging in some practical testing. In humans as in crows, it seems that performance on physical tasks may not always be based on a strong form of a priori understanding, but that understanding may in fact be promoted by some practical engagement with the task itself (a similar point was made by Hunt, Rutledge, et al., 2006). Concluding Remarks Betty’s manufacture of hooks from novel materials and Guillem’s performance in the modified trap-tube task are, to our knowledge, amongst the closest any non-humans have come to demonstrating an ‘understanding’ of folk physics. However, as we have mentioned earlier, the concept of understanding is an epistemological minefield, and in what follows we examine at greater length some of the issues raised by this word and their implications for comparative cognition research. In the earlier description of Betty’s wire-bending behaviour, we stated that "she was able to invent a method of shaping the material into a functional and probably preconceived form." If ‘invent’ were exchanged for ‘discover’ and ‘preconceived’ were swapped for ‘previously learned,’ there would be no need to look beyond associative learning and generalisation, albeit of a relatively complex and unlikely nature. We chose the original wording partly as a teaser (surely some readers’ reservations about anthropomorphic language must have been aroused by the sentence) and partly because, in our opinion, this possibility is worth entertaining as a working hypothesis. We adhere to the spirit of Morgan’s Canon (Thomas, 2001), and certainly do not argue that, because Betty acted as a human might, she did so on the same conceptual basis (‘argument by analogy’; see Povinelli, Bering, & Giambrone, 2000). However, to dismiss the possibility of higher cognition out-of-hand could be construed as anthropocentric in itself (Keeley, 2004). On the basis of the available evidence, we cannot yet exclude the possibility that all apparently intelligent behaviours shown by New Caledonian crows can be explained by associative learning mechanisms, and the same may be true for other non-human species. Given this situation, is it still meaningful or useful to refer to understanding in crows and in animal behaviour in general? The term ‘understanding’ has been discussed in great depth by philosophers (e.g., Overskeid, 2005; Searle, 1980), and there is still no general agreement about how it should be defined or how it can be conclusively demonstrated, even in humans. We have mentioned Overkeid’s definition that "understanding X means not believing lack of knowledge about X to be an obstacle to reaching a relevant, currently active goal" (Overskeid, 2005, p. 601), which implies that understanding is a default state, and most animals (including people) "are probably in a state of understanding most of the time" (p. 612). Overskeid generated this definition by analysing situations in which humans would claim that they did or did not understand something, a process which—while entirely valid from a philosophical point of view—necessarily produces a definition from a first-person perspective that is unsuitable for application in animal cognition research. Overskeid proposes that we can judge understanding in others by observing whether they behave appropriately in certain situations; if so, we can ascribe to them some degree of understanding in the relevant domain. For example, if someone anticipates rain and prepares herself by carrying an umbrella, Overskeid would infer that she had some understanding of meteorology. However, this definition takes no account of the process by which the subject acquired proficiency in that situation—if, for example, the subject evolved in an environment where the weather could be predicted by simple, reliable cues, and she was genetically endowed with rules that enabled accurate weather prediction, it would seem inappropriate to ascribe any degree of understanding to her, regardless of her successful performance (indeed, plants ‘predict’ the weather every spring when the days get longer). This definition also says nothing about what the subject understands or the correctness of her understanding. In the domain of folk physics, when we say that an animal ‘understands’ the problem, we usually intend to mean that they have some (more-or-less correct) knowledge of the causal basis of the task (Vonk, 2005). However, this introduces new problems with what we mean by ‘knowledge’ and ‘causal basis.’ Knowledge can be obtained in various ways, one of which is associative learning, and there is no universal agreement on how to separate associatively-learned knowledge from associatively-learned behaviour. Equally, most humans have only a limited, proximate understanding of the causal basis of phenomena around them, including their actions. For instance, one can say that the reason images disappeared from the TV screen is because the set was unplugged, but this does not imply the converse: that the subject understands why and how plugging the TV in causes the images to be formed. Thus, a pertinent question is how accurate and detailed causal understanding needs to be for us to recognise it as such. We do not have a precise answer and remain in the quandary described in the introduction: At the moment there is no definite test for understanding. We believe, however, that progress is possible if one relaxes the epistemological strictures just enough to foster empirical research, while avoiding over-interpreting results. The role of theory in this case resembles that of evolutionary assumptions in much of behavioural ecology: While precise definitions of fitness remain a matter of debate (Grafen, 1999), predictive models based on intuitive components of fitness, such as foraging performance, continue to lead to significant advances. Our brief discussion of philosophical matters testifies to our interest in the wider implications of our research. New Caledonian crows and other corvids exhibit behaviour that looks ‘smart’ to most human observers, and some observations cry out for comparative work with other seemingly intelligent animals, notably humans and other primates. Such comparisons are important and have produced some striking examples of convergent evolution in birds and mammals. Empirical evidence for corvid intelligence is accumulating rapidly and has led some authors to propose that "corvids should perhaps be considered as ‘feathered’ apes" (Emery, 2004, p. 182; see also Clayton & Emery, 2005). We naturally share the general enthusiasm for corvid research but think that care needs to be taken when interpreting and describing results. In particular, we need to be cautious of ascribing general cognitive abilities to the whole Corvidae family based on the abilities of individual species, which may reflect adaptations to specific ecological niches. For example, western scrub-jays and common ravens rely heavily on food caching and raiding conspecifics’ caches, and elegant experimental investigations have demonstrated that they are equipped with complex and flexible cache-protection strategies and mechanisms (e.g., Bugnyar & Heinrich, 2005; Dally, Emery, & Clayton, 2005, 2006; Emery & Clayton, 2001). Similarly, pinyon jays (Gymnorhinus cyanocephalus) are highly social, and can use transitive reasoning to predict the dominance of conspecifics (Paz-y-Miño C., Bond, Kamil, & Balda, 2004). Whilst it is possible that these abilities are equivalent to the similar cognitive processes in humans and are shared by the whole Corvidae family, it could equally be the case that they are species-specific adaptations. The remarkable tool-oriented behaviour of New Caledonian crows may be another example of such an adaptation. In contrast, many of these cognitive abilities appear to be found in all species of great ape (Tomasello & Call, 1997) and are exhibited in a wide variety of different contexts, making it more plausible that in apes, they are truly the consequence of some kind of ‘general’ intelligence. However, in many ways corvid research is still decades behind primate research, particularly in terms of long-term field studies that are necessary for putting experimental findings into an ecological and evolutionary perspective, so it would be premature to reach firm conclusions at this stage (a point also made by others, e.g., Emery, 2006; Emery & Clayton, 2004a). We have recently attempted to identify possible evolutionary origins of the behavioural action patterns that lead to tool use in New Caledonian crows (Kenward et al., 2007). Thomas Bugnyar and co-workers from Austria had documented in detail the development of juvenile common ravens, a species that does not habitually use tools in the wild, but is otherwise renowned for its cognitive capacities (Bugnyar & Heinrich, 2005; Heinrich, 1999). Taking advantage of the fact that the Austrian and the Oxford research groups had employed similar (albeit not identical) observation protocols for their longitudinal studies, we compared the ontogeny of object-oriented behaviours between the two species. Our analyses revealed striking developmental similarities between TOB in crows and food-caching behaviour in ravens, including similar precursor behaviours. Given that the common ancestor of New Caledonian crows and ravens was almost certainly a caching species (de Kort & Clayton, 2006), we hypothesise that the action patterns for tool use in crows have their evolutionary origins in caching behaviour. Tool use in New Caledonian crows is the result of natural selection, yet we remain largely ignorant of the selective forces that may have fostered this unusual behaviour in the evolutionary past and those that maintain it under present-day conditions. In 2005 we launched a long-term research project to study New Caledonian crows in the wild. It is our hope that our observations in the field, together with those of our colleagues from New Zealand, will inform experimental work with captive crows. This concerted work in the lab and in the field may yield answers to the many questions we were unable to address in this review. References Alcock, J. (1972). The evolution of the use of tools by feeding animals. Evolution, 26, 464-473. Amsel, E., Goodman, G., Savoie, D., & Clark, M. (1996). The development of reasoning about causal and noncausal influences on levers. Child Development, 67, 1624-1646. Anderson, J. R., & Henneman, M. C. (1994). Solutions to a tool-use problem in a pair of Cebus apella. Mammalia, 58, 351-361. Andersson, S. (1989). Tool use by the fan-tailed raven (Corvus rhipidurus). Condor, 91, 999. Barnard, C. J. (2004). Animal behaviour: Mechanism, development, function and evolution. Harlow: Prentice Hall. Beck, B. B. (1980). Animal tool behavior: The use and manufacture of tools by animals. New York, NY: Garland. Beck, B. B. (1986). Tools and intelligence. In R. J. Hoage & L. Goldman (Eds.), Animal intelligence: Insights into the animal mind (pp. 135-147). Washington D.C.: Smithsonian Institution Press. Biro, D., Inoue-Nakamura, N., Tonooka, R., Yamakoshi, G., Sousa, C., & Matsuzawa, T. (2003). Cultural innovation and transmission of tool use in wild chimpanzees: Evidence from field experiments. Animal Cognition, 6, 213-223. Blaisdell, A. P., Sawa, K., Leising, K. J., & Waldmann, M. R. (2006). Causal reasoning in rats. Science, 311, 1020-1022. Boesch, C., & Boesch, H. (1990). Tool use and tool making in wild chimpanzees. Folia Primatologica, 54, 86-99. Boesch, C., & Boesch, H. (1993). Diversity of tool use and tool-making in wild chimpanzees. In A. Berthelet & J. Chevaillon (Eds.), The use of tools by human and non-human primates (pp. 158-187). Oxford: Clarendon Press. Borsari, A., & Ottoni, E. B. (2005). Preliminary observations of tool use in captive hyacinth macaws (Anodorhynchus hyacinthinus). Animal Cognition, 8, 48-52. Boswall, J. (1978). Further notes on tool using by birds and related behaviour. Avicultural Magazine, 84, 162-166. Boswall, J. (1983). Tool-using and related behaviour by birds: More notes. Avicultural Magazine, 89, 94-108. Boyd, R., & Silk, J. B. (2006). How humans evolved (4th ed.). New York: W.W. Norton. Bugnyar, T., & Heinrich, B. (2005). Ravens, Corvus corax, differentiate between knowledgeable and ignorant competitors. Proceedings of the Royal Society of London B, 272, 1641-1646. Caffrey, C. (2000). Tool modification and use by an American Crow. Wilson Bulletin, 112, 283-284. Call, J. (2004). Inferences about the location of food in the great apes (Pan paniscus, Pan troglodytes, Gorilla gorilla, and Pongo pygmaeus). Journal of Comparative Psychology, 118, 232-241. Chappell, J. (2006). Avian cognition: Understanding tool use. Current Biology, 16, R244-R245. Chappell, J., & Kacelnik, A. (2002). Tool selectivity in a non-mammal, the New Caledonian crow (Corvus moneduloides). Animal Cognition, 5, 71-78. Chappell, J., & Kacelnik, A. (2004). Selection of tool diameter by New Caledonian crows Corvus moneduloides. Animal Cognition, 7, 121-127. Chappell, J., & Kacelnik, A. (2007). Folk physics in New Caledonian crows: Do crows understand gravity? Manuscript in preparation. Clayton, N. S. (1992). The ontogeny of food-storing and retrieval in marsh tits. Behaviour, 122, 11-25. Clayton, N. S., & Emery, N. J. (2005). Corvid cognition. Current Biology, 15, R80-R81. Cole, P. D. (2004). The ontogenesis of innovative tool use in an American Crow (Corvus brachyrhynchos). Unpublished doctoral dissertation, Dalhousie University, Canada, Halifax, Nova Scotia. Collias, E. C., & Collias, N. E. (1964). The development of nest-building behaviour in a weaverbird. Auk, 81, 42-52. Cristol, D. A., & Switzer, P. V. (1999). Avian prey-dropping behavior. II. American crows and walnuts. Behavioral Ecology, 10, 220-226. Dally, J. M., Emery, N. J., & Clayton, N. S. (2005). Cache protection strategies by western scrub-jays, Aphelocoma californica: Implications for social cognition. Animal Behaviour, 70, 1251-1263. Dally, J. M., Emery, N. J., & Clayton, N. S. (2006). Food-caching Western scrub-jays keep track of who was watching when. Science, 312, 1662-1665. de Houwer, J., Beckers, T., & Vandorpe, S. (2005). Evidence for the role of higher order reasoning processes in cue competition and other learning phenomena. Learning & Behavior, 33, 239-249. de Kort, S. R., & Clayton, N. S. (2006). An evolutionary perspective on caching by corvids. Proceedings of the Royal Society of London B, 273, 417-423. Délacour, J. (1966). Guide des oiseaux de la Nouvelle-Calédonie et de ses dépendances. Paris: Delachaux et Niestlé. Dickinson, A. (2001). Causal learning: Association versus computation. Current Directions in Psychological Science, 10, 127-132. Emery, N. J. (2004). Are corvids ‘feathered apes’? Cognitive evolution in crows, jays, rooks and jackdaws. In S. Watanabe (Ed.), Comparative analysis of minds (pp. 181-213). Tokyo: Keio University Press. Emery, N. J. (2006). Cognitive ornithology: The evolution of avian intelligence. Philosophical Transactions of the Royal Society of London B, 361, 23-43. Emery, N. J., & Clayton, N. S. (2001). Effects of experience and social context on prospective caching strategies by scrub jays. Nature, 414, 443-446. Emery, N. J., & Clayton, N. S. (2004a). Comparing the complex cognition of birds and primates. In L. J. Rogers & G. Kaplan (Eds.), Comparative vertebrate cognition: Are primates superior to non-primates? (pp. 3-55). New York: Kluwer Academic / Plenum Publishers. Emery, N. J., & Clayton, N. S. (2004b). The mentality of crows: Convergent evolution of intelligence in corvids and apes. Science, 306, 1903-1907. Fitch, W. T., Hauser, M. D., & Chomsky, N. (2005). The evolution of the language faculty: Clarifications and implications. Cognition, 97, 179-210. Fragaszy, D. M., & Adams-Curtis, L. E. (1997). Developmental changes in manipulation in tufted capuchins (Cebus apella) from birth through 2 years and their relation to foraging and weaning. Journal of Comparative Psychology, 111, 201-211. Freeman, S. (2002). Biological science (2nd ed.). Upper Saddle River, NJ: Pearson Prentice Hall. Galef, B. G., Jr. (1988). Imitation in animals: History, definition, and interpretation of data from the psychological laboratory. In T. R. Zentall & B. G. Galef, Jr (Eds.), Social learning: Psychological and biological perspectives (pp. 3-28). Hillsdale, NJ: Lawrence Erlbaum Associates, Inc. Gayou, D. C. (1982). Tool use by green jays. Wilson Bulletin, 94, 593-594. Gibson, E. J., & Pick, A. D. (2000). An ecological approach to perceptual learning and development. Oxford: Oxford University Press. Grafen, A. (1999). Formal Darwinism: The individual-as-maximizing-agent analogy and bet-hedging. Proceedings of the Royal Society of London B, 266, 799-803. Hall, K. R. L. (1963). Tool-using performances as indicators of behavioural adaptability. Current Anthropology, 4, 479-494. Hansell, M. H. (1984). Animal architecture and building behaviour. London: Longman. Hansell, M. H. (1987, January 8). What’s so special about using tools? New Scientist, 1542, 55-56. Hansell, M. H. (2005). Animal architecture. Oxford: Oxford University Press. Hauser, M. D. (1997). Artifactual kinds and functional design features: What a primate understands without language. Cognition, 64, 285-308. Hauser, M. D., Kralik, J., & Botto-Mahan, C. (1999). Problem solving and functional design features: Experiments on cotton-top tamarins, Saguinus oedipus oedipus. Animal Behaviour, 57, 565-582. Hauser, M. D., Pearson, H., & Seelig, D. (2002). Ontogeny of tool use in cottontop tamarins, Saguinus oedipus: Innate recognition of functionally relevant features. Animal Behaviour, 64, 299-311. Hauser, M. D., & Santos, L. R. (in press). The evolutionary ancestry of our knowledge of tools: From percepts to concepts. In E. Margolis & S. Lawrence (Eds.), Creations of the mind. Oxford, UK: Oxford University Press. Hauser, M. D., Santos, L. R., Spaepen, G. M., & Pearson, H. E. (2002). Problem solving, inhibition and domain-specific experience: Experiments on cottontop tamarins, Saguinus oedipus. Animal Behaviour, 64, 387-396. Hayashi, M., & Matsuzawa, T. (2003). Cognitive development in object manipulation by infant chimpanzees. Animal Cognition, 6, 225-233. Heinrich, B. (1999). The mind of the raven: Investigations and adventures with wolf-birds. New York: Cliff Street Books. Henty, C. J. (1986). Development of snail-smashing by song thrushes. British Birds, 79, 277-281. Heyes, C. M. (1994). Social learning in animals: Categories and mechanisms. Biological Reviews of the Cambridge Philosophical Society, 69, 207-231. Hicks, T. C., Fouts, R. S., & Fouts, D. H. (2005). Chimpanzee (Pan troglodytes troglodytes) tool use in the Ngotto Forest, Central African Republic. American Journal of Primatology, 65, 221-237. Hunt, G. R. (1996). Manufacture and use of hook-tools by New Caledonian crows. Nature, 379, 249-251. Hunt, G. R. (2000a). Human-like, population-level specialization in the manufacture of pandanus tools by New Caledonian crows Corvus moneduloides. Proceedings of the Royal Society of London B, 267, 403-413. Hunt, G. R. (2000b). Tool use by the New Caledonian crow Corvus moneduloides to obtain Cerambycidae from dead wood. Emu, 100, 109-114. Hunt, G. R., Corballis, M. C., & Gray, R. D. (2001). Laterality in tool manufacture by crows. Nature, 414, 707. Hunt, G. R., Corballis, M. C., & Gray, R. D. (2006). Design complexity and strength of laterality are correlated in New Caledonian crows’ pandanus tool manufacture. Proceedings of the Royal Society of London B, 273, 1127-1133. Hunt, G. R., & Gray, R. D. (2002). Species-wide manufacture of stick-type tools by New Caledonian Crows. Emu, 102, 349-353. Hunt, G. R., & Gray, R. D. (2003). Diversification and cumulative evolution in New Caledonian crow tool manufacture. Proceedings of the Royal Society of London B, 270, 867-874. Hunt, G. R., & Gray, R. D. (2004a). The crafting of hook tools by wild New Caledonian crows. Proceedings of the Royal Society of London B (Suppl.), 271, S88-S90. Hunt, G. R., & Gray, R. D. (2004b). Direct observations of pandanus-tool manufacture and use by a New Caledonian crow (Corvus moneduloides). Animal Cognition, 7, 114-120. Hunt, G. R., Rutledge, R. B., & Gray, R. D. (2006). The right tool for the job: What strategy do wild New Caledonian crows use? Animal Cognition, 9, 307-316. Hunt, G. R., Sakuma, F., & Shibata, Y. (2002). New Caledonian crows drop candle-nuts onto rock from communally-used forks on branches. Emu, 102, 283-290. Jones, T., & Kamil, A. C. (1973). Tool-making and tool-using in the northern blue jay. Science, 180, 1076-1078. Kacelnik, A., Chappell, J., Weir, A. A. S., & Kenward, B. (2006). Cognitive adaptations for tool-related behaviour in New Caledonian crows. In E. A. Wasserman & T. R. Zentall (Eds.), Comparative cognition: Experimental explorations of animal intelligence (pp. 515-528). Oxford: Oxford University Press. Keeley, B. L. (2004). Anthropomorphism, primatomorphism, mammalomorphism: Understanding cross-species comparisons. Biology & Philosophy, 19, 521-540. Kenward, B. (2006). Why do New Caledonian crows use tools? Unpublished doctoral dissertation, University of Oxford, Oxford. Kenward, B., Rutz, C., Weir, A. A. S., Chappell, J., & Kacelnik, A. (2004). Morphology and sexual dimorphism of the New Caledonian Crow Corvus moneduloides, with notes on its behaviour and ecology. Ibis, 146, 652-660. Kenward, B., Rutz, C., Weir, A. A. S., & Kacelnik, A. (2006). Development of tool use in New Caledonian crows: Inherited action patterns and social influence. Animal Behaviour, 72, 1329-1343. Kenward, B., Schlögl, C., Rutz, C., Weir, A. A. S., Bugnyar, T., & Kacelnik, A. (2007). On the evolutionary origin of tool-oriented behaviour in New Caledonian crows. Manuscript in preparation. Kenward, B., Weir, A. A. S., Rutz, C., & Kacelnik, A. (2005). Tool manufacture by naive juvenile crows. Nature, 433, 121. Laland, K. N., & Janik, V. M. (2006). The animal cultures debate. Trends in Ecology & Evolution, 21, 542-547. Layard, E. L., & Layard, E. L. C. (1882). Notes on the avifauna of New Caledonia. Ibis, 6, 520-522. Lefebvre, L., Reader, S. M., & Sol, D. (2004). Brains, innovations and evolution in birds and primates. Brain, Behavior and Evolution, 63, 233-246. Limongelli, L., Boysen, S. T., & Visalberghi, E. (1995). Comprehension of cause-effect relations in a tool-using task by chimpanzees (Pan troglodytes). Journal of Comparative Psychology, 109, 18-26. Lockman, J. J. (2000). A perception-action perspective on tool use development. Child Development, 71, 137-144. Lonsdorf, E. V. (2005). Sex differences in the development of termite-fishing skills in the wild chimpanzees, Pan troglodytes schweinfurthii, of Gombe National Park, Tanzania. Animal Behaviour, 70, 673-683. Lonsdorf, E. V. (2006). What is the role of mothers in the acquisition of termite-fishing behaviors in wild chimpanzees (Pan troglodytes schweinfurthii)? Animal Cognition, 9, 36-46. Machado, A., & Silva, F. J. (2003). You can lead an ape to a tool, but …: A review of Povinelli’s Folk physics for apes: The chimpanzee’s theory of how the world works. Journal of the Experimental Analysis of Behavior, 79, 267-286. Marler, P., & Slabbekoorn, H. (Eds.). (2004). Nature’s music: The science of birdsong. Amsterdam: Elsevier Academic Press. Mulcahy, N. J., Call, J., & Dunbar, R. I. M. (2005). Gorillas (Gorilla gorilla) and orangutans (Pongo pygmaeus) encode relevant problem features in a tool-using task. Journal of Comparative Psychology, 119, 23-32. Neuringer, A., Kornell, N., & Olufs, M. (2001). Stability and variability in extinction. Journal of Experimental Psychology: Animal Behavior Processes, 27, 79-94. Nishida, T. (1972). The ant-gathering behaviour by the use of tools among wild chimpanzees of the Mahali Mountains. Journal of Human Evolution, 2, 357-370. Orenstein, R. (1972). Tool use by the New Caledonian crow (Corvus moneduloides). Auk, 89, 674-676. Overskeid, G. (2005). Empirically understanding understanding can make problems go away: The case of the Chinese room. Psychological Record, 55, 595-617. Parker, S. T., & Gibson, K. R. (1977). Object manipulation, tool use and sensorimotor intelligence as feeding adaptations in Cebus monkeys and great apes. Journal of Human Evolution, 6, 623-641. Paz-y-Miño C., G., Bond, A. B., Kamil, A. C., & Balda, R. P. (2004). Pinyon jays use transitive inference to predict social dominance. Nature, 430, 778-781. Penn, D. C., & Povinelli, D. J. (2007). Causal cognition in human and nonhuman animals: A comparative, critical review. Annual Review of Psychology, 58, 97-118. Pouydebat, E., Berge, C., Gorce, P., & Coppens, Y. (2005). Use and manufacture of tools to extract food by captive Gorilla gorilla gorilla: Experimental approach. Folia Primatologica, 76, 180-183. Povinelli, D. J. (Ed.). (2000). Folk physics for apes. Oxford, UK: Oxford University Press. Povinelli, D. J., Bering, J. M., & Giambrone, S. (2000). Toward a science of other minds: Escaping the argument by analogy. Cognitive Science, 24, 509-541. Povinelli, D. J., Reaux, J. E., Theall, L. A., & Giambrone, S. (2000a). The question of tool modification. In D. J. Povinelli (Ed.), Folk physics for apes (pp. 271-296). Oxford, UK: Oxford University Press. Povinelli, D. J., Reaux, J. E., Theall, L. A., & Giambrone, S. (2000b). The rope, hook, touching-stick, and related problems: The question of physical connection. In D. J. Povinelli (Ed.), Folk physics for apes (pp. 206-253). Oxford, UK: Oxford University Press. Reader, S. M., Sol, D., & Lefebvre, L. (2005). Comparing cognition across species. Trends in Cognitive Sciences, 9, 411. Reaux, J. E., & Povinelli, D. J. (2000). The trap-tube problem. In D. J. Povinelli (Ed.), Folk physics for apes (pp. 108-131). Oxford, UK: Oxford University Press. Reid, J. B. (1982). Tool-use by a rook (Corvus frugilegus), and its causation. Animal Behaviour, 30, 1212-1216. Roth, G., & Dicke, U. (2005). Evolution of the brain and intelligence. Trends in Cognitive Sciences, 9, 250-257. Rutledge, R., & Hunt, G. R. (2004). Lateralized tool use in wild New Caledonian crows. Animal Behaviour, 67, 327-332. Santos, L. R., Miller, C. T., & Hauser, M. D. (2003). Representing tools: How two non-human primate species distinguish between the functionally relevant and irrelevant features of a tool. Animal Cognition, 6, 269-281. Santos, L. R., Pearson, H., Spaepen, G. M., Tsao, F., & Hauser, M. (2006). Probing the limits of tool competence: Experiments with two non-tool-using species (Cercopithecus aethiops and Saguinus oedipus). Animal Cognition, 9, 94-109. Santos, L. R., Rosati, A., Sproul, C., Spaulding, B., & Hauser, M. D. (2005). Means-means-end tool choice in cotton-top tamarins (Saguinus oedipus): Finding the limits on primates’ knowledge of tools. Animal Cognition, 8, 236-246. Sarsby, J. (Writer). (1998). The insatiable appetite [Television series episode]. In M. Salisbury (Producer), The life of birds. UK: BBC Natural History Unit. Searle, J. R. (1980). Minds, brains, and programs. Behavioral and Brain Sciences, 3, 417-425. Seed, A. M., Tebbich, S., Emery, N. J., & Clayton, N. S. (2006). Investigating physical cognition in rooks, Corvus frugilegus. Current Biology, 16, 697-701. Silva, F. J., Page, D. M., & Silva, K. M. (2005). Methodological-conceptual problems in the study of chimpanzees’ folk physics: How studies with adult humans can help. Learning & Behavior, 33, 47-58. Spaulding, B., & Hauser, M. D. (2005). What experience is required for acquiring tool competence? Experiments with two Callitrichids. Animal Behaviour, 70, 517-526. Tebbich, S., & Bshary, R. (2004). Cognitive abilities related to tool use in the woodpecker finch, Cactospiza pallida. Animal Behaviour, 67, 689-697. Tebbich, S., Seed, A. M., Emery, N. J., & Clayton, N. S. (in press). Non-tool-using rooks (Corvus frigilegus) solve the trap-tube task. Animal Cognition. Tebbich, S., Taborsky, M., Fessl, B., & Blomqvist, D. (2001). Do woodpecker finches acquire tool-use by social learning? Proceedings of the Royal Society of London B, 268, 2189-2193. Tebbich, S., Taborsky, M., Fessl, B., Dvorak, M., & Winkler, H. (2004). Feeding behavior of four arboreal Darwin’s finches: Adaptations to spatial and seasonal variability. Condor, 106, 95-105. Thomas, R. K. (2001). Lloyd Morgan’s Canon: A history of misrepresentation. History & Theory of Psychology Eprint Archive, 2006. Retrieved 26 September, 2006, from http://htpprints.yorku.ca/archive/00000017/ Thorndike, E. L. (1898). Animal intelligence: An experimental study of the associative processes in animals. Psychological Monographs, 2, 1-109. Thouless, C. R., Fanshawe, J. H., & Bertram, B. C. R. (1989). Egyptian vultures Neophron percnopterus and ostrich Struthio camelus eggs: The origins of stone-throwing behavior. Ibis, 131, 9-15. Tomasello, M., & Call, J. (1997). Primate cognition. New York, NY: Oxford University Press. Townsend, M. (2005, March 6). Sheep might be dumb ... but they’re not stupid. The Observer, p. 10. Ueno, Y., & Fujita, K. (1998). Spontaneous tool use by a Tonkean macaque (Macaca tonkeana). Folia Primatologica, 69, 318-324. Vargha-Khadem, F., Gadian, D. G., Copp, A., & Mishkin, M. (2005). FOXP2 and the neuroanatomy of speech and language. Nature Reviews Neuroscience, 6, 131-138. Visalberghi, E. (1993). Tool use in a South American monkey species: An overview of the characteristics and limits of tool use in Cebus apella. In A. Berthelet & J. Chevaillon (Eds.), The use of tools by human and non-human primates (pp. 118-135). Oxford: Clarendon Press. Visalberghi, E. (1997). Success and understanding in cognitive tasks: A comparison between Cebus apella and Pan troglodytes. International Journal of Primatology, 18, 811-830. Visalberghi, E., Fragaszy, D. M., & Savage-Rumbaugh, E. S. (1995). Performance in a tool-using task by common chimpanzees (Pan troglodytes), bonobos (Pan paniscus), an orangutan (Pongo pygmaeus), and capuchin monkeys (Cebus apella). Journal of Comparative Psychology, 109, 52-60. Visalberghi, E., & Limongelli, L. (1994). Lack of comprehension of cause-effect relations in tool-using capuchin monkeys (Cebus apella). Journal of Comparative Psychology, 108, 15-22. Visalberghi, E., & Trinca, L. (1989). Tool use in capuchin monkeys: Distinguishing between performing and understanding. Primates, 30, 511-521. Vonk, J. (2005). Causality in non-humans. In A. Reboul & G. Origgi (Eds.), Causality. Institute for Cognitive Science and the University of Geneva. Weir, A. A. S. (2006). Cognitive psychology of tool use in New Caledonian crows (Corvus moneduloides). Unpublished doctoral dissertation, University of Oxford, Oxford. Weir, A. A. S., Chappell, J., & Kacelnik, A. (2002). Shaping of hooks in New Caledonian crows. Science, 297, 981. Weir, A. A. S., & Kacelnik, A. (2006). A New Caledonian crow (Corvus moneduloides) creatively re-designs tools by bending or unbending aluminium strips. Animal Cognition, 9, 317-334. Wolpert, L. (2003). Causal belief and the origins of technology. Philosophical Transactions of the Royal Society of London A, 361, 1709 -1719. |







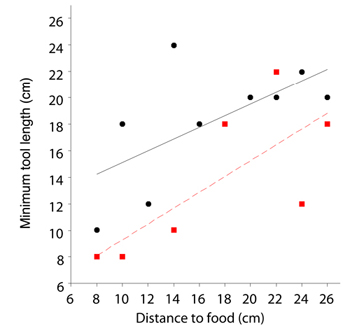
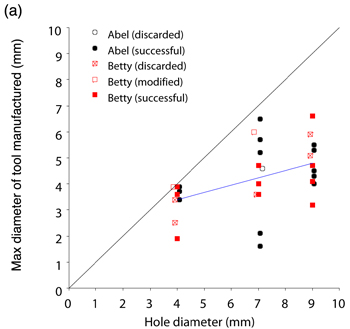
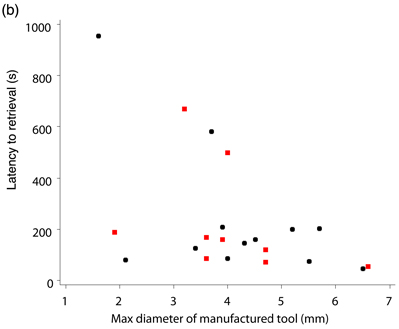
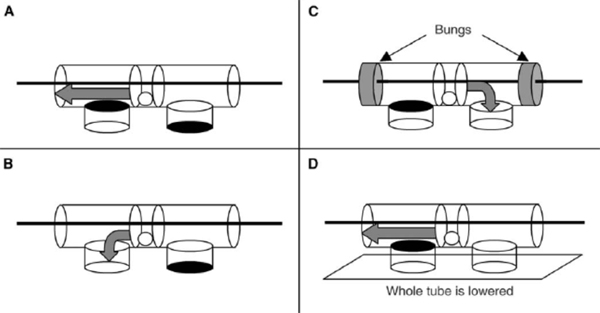



Lucas A. Bluff, Alex A. S. Weir, Christian Rutz, Joanna H. Wimpenny, and Alex Kacelnik, Department of Zoology, University of Oxford, Oxford, UK.
|