| <<Vol. 2 Table of Contents | |
|
|
|
|
2007 |
Volume 2, pp 79-92 |

Issues in the Comparative Cognition of Abstract-Concept Learning
Jeffrey S. Katz
Auburn University
Anthony A. Wright
University of Texas Health Science Center at Houston
Kent D. Bodily
Auburn University
|
|
||||||||||||||||||||||
|
||||||||||||||||||||||
|
Abstract concepts are said to be the basis of higher order cognition in human, and no concept is more important than the concept of identity. William James (1890/1950) was perhaps the first to note that our "sense of sameness is the very keel and backbone of our thinking." (p. 459). Over the past century, the ability to judge whether items are the same or different has been a central focus in cognitive development, cognition, and comparative cognition (e.g., Daehler & Bukatko, 1985; Mackintosh, 2000; Shettleworth, 1998; Thompson & Oden, 2000). For example, abstract thinking is considered to be the basis of equivalence operations in math (e.g., "item I is the same as item J"), conservation tasks, and may be a necessary prerequisite for learning language (e.g., Marcus et al., 1999; Piaget & Inhelder, 1966/1969; Siegler, 1996). This thinking is abstract because it is based on rules which allow subjects’ judgments to transcend the training stimuli and is therefore called higher order. Abstract-concept learning is the focus of this article. Issues of testing and verifying abstract-concept learning with animals in same/different (S/D) and matching-to-sample (MTS) tasks are discussed. In addition, we review some of the research on abstract-concept learning. In cognitive and comparative psychology, abstract-concept learning is, unfortunately, often confused with categorization because both are frequently referred to as concept learning. To clarify, there are two types of categorization, 1) natural concepts, which are accounted for by stimulus generalization based on specific features, and 2) associative concepts, which are accounted for by second-order conditioning. Natural concept learning (also called perceptual concept learning) involves categorizing (sorting) stimuli (e.g., those found in nature like pictures of birds, flowers, people, or artificial ones like shapes) based on stimulus perceptual similarity into appropriate categories (e.g., Herrnstein, Loveland, & Cable 1976; Medin, 1989; Wasserman, Kiedinger, & Bhatt, 1988). Associative concept learning (also called functional concept learning) involves categorizing stimuli based on a common response or outcome regardless of perceptual similarity into appropriate categories (e.g., Urcuioli, 2006; Vaughan, 1988; Wasserman, DeVolder, & Coppage, 1992). Abstract-concept learning is different. Abstract-concept learning cannot be accounted for by these generalization processes. Abstract-concept learning involves judging a relationship between stimuli based on a rule (e.g., identity, difference, oddity, greater than, addition, subtraction). The rule is considered to be abstract when it can be applied to novel stimuli. Since Thorndike’s (1911/1998) experimental studies in comparative psychology, theories of animal intelligence have focused on cross-species comparisons in their ability to learn abstract concepts (e.g., D’Amato, Salmon, & Colombo 1985; Mackintosh, 1988; Premack, 1983b; Thompson, 1995). Abstract-concept learning has been the centerpiece of theories of animal intelligence because it represents higher-order learning. In terms of hierarchies of learning abilities, categorization is considered to be a lower level of learning than abstract-concept learning. Many theories of animal intelligence have been centered on levels of learning (e.g., Herrnstein, 1990; Thomas, 1980, 1996; Thompson & Oden, 2000). As a result, nonhuman animals believed to form abstract concepts were thought to be more intelligent than those presumed to not reach the higher level (e.g., Premack, 1978, 1983a). The focus on hierarchies of learning abilities has produced an emphasis on discovering which species (i.e., who) have or do not have a particular ability. This approach is akin to the cognitive-modular approach, which argues that failure to pass a test (to attain a higher level) is evidence that a species lacks the requisite cognitive module to do so (cf., Tooby & Cosmides, 2000). Consequently, this focus on who has resulted in single experiments, often with limited parameter manipulation, to demonstrate whether or not a species has the ability to learn abstract concepts (generally) and thereby determine if it has inherited the requisite cognitive module. In essence, this endeavor boils down to a search for qualitative differences. The problem with such an all-or-none approach is that one assumes that the procedures adequately assessed the abilities of each species. Some procedures do not adequately fit the predispositions of the species and can create what appear to be qualitative species differences based on, for example, the level of transfer in abstract-concept learning tasks. The approach is tantamount to proving that transfer failure is a cognitive inadequacy not a procedural inadequacy. The problem faced by experimenters is to discover procedures that are adequate for testing abstract-concept learning. Hence, it becomes difficult to conduct the task variations (called systematic variation) to insure that some alleged cognitive inability is really lacking (but see Bitterman, 1965, 1975; Kamil, 1988). That is, to compare the functional relationship of some variable across species one needs to know whether performance depends on the same variable for the species in question. If the answer is unknown, then judgment should be suspended. Doing otherwise is trying to prove the null hypothesis (Macphail, 1985). A different approach to abstract-concept learning is the general-process account. There are, to be sure, degrees of generality. A strong version of the general-process view would be that all vertebrates have the abstract-concept learning ability, regardless of whether the ability evolved through their homology or homoplasy. A somewhat weaker version of the general-process view would be that a wide variety of animals (e.g., food storing birds) have the ability (cf. Papini, 2002). For sure, there will be differences in abstract-concept learning abilities across species, but the key, in our view, is to discover whether or not there are conditions that reveal the generality in their eventual ability to fully learn an abstract concept. Generality can be revealed by exploring how abstract-concept learning works under various parameter manipulations. This approach can ultimately reveal whether a difference in performance across species is a qualitative difference (i.e., the presence or absence of a cognitive module) or a quantitative difference (i.e., a general process). A focus on how instead of who should be productive in discovering the mechanisms that underlie abstract-concept learning in a variety of evolutionarily diverse species. This approach to studying abstract-concept learning is supported by positive transfer results from phylogenetically diverse species which were once thought to be incapable of S/D abstract-concept learning (i.e., a qualitative absence in this cognitive module), including language-naïve chimpanzees (Thompson, Oden, & Boysen, 1997), baboons (Bovet & Vauclair, 2001), capuchin monkeys (Wright, Rivera, Katz, & Bachevalier, 2003), rhesus monkeys (Wright, Santiago, Urcuioli, & Sands, 1984), parrots (Pepperberg, 1987), and pigeons (Katz & Wright, 2006). Additional studies have found evidence consistent with S/D abstract-concept learning, however, due to procedural limitations, the results are open to alternative explanations. The issues we discuss next center around the criteria that we believe are important to achieve in order to rule out alternative explanations. Importance of Abstract-Concept Learning Criteria Over the past 30 years the criteria for abstract-concept learning has become a moving target (Cook 2002; Premack, 1978; Wright, 1991; Wright & Katz, 2006). As new findings have been revealed new requirements have been imposed. In this section we present criteria that we and others believe are important to establish abstract-concept learning because they can help rule out alternative explanations based on the novelty of the stimuli (criteria 1 and 2) and inconclusive results (criterion 3).
Importance of Two-Item Same/Different Discrimination
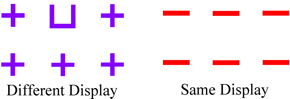
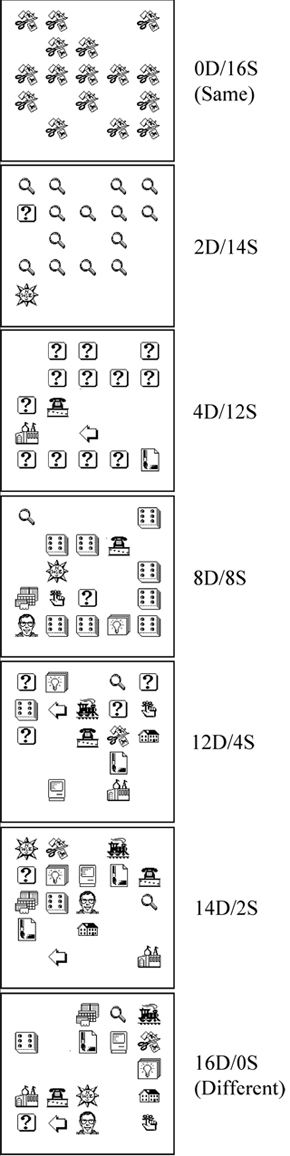
Another issue, specific to conclusions concerning abstract same/different concept learning, is the use of two-items in a stimulus display. Any species that is purported to be able to learn a S/D concept should be able to do it with two items. When we developed our procedure for pigeons and first presented our findings at the International Conference on Comparative Cognition in 2001, only one published study had found evidence that pigeons could learn an abstract S/D concept with two-item displays to any high degree of accuracy (72%; Santiago & Wright, 1984). Since that time other articles have been published showing some support for two-item S/D concept learning in pigeons (Blaisdell & Cook, 2005; Cook, Kelly, & Katz, 2003). Like the Santiago & Wright (1984) study both of these studies found partial transfer. Partial transfer, as opposed to full transfer, is suggestive and encouraging but is inconclusive due to the possibility that multiple cues are controlling behavior (criterion 3). The interpretation of the results from Blaisdell & Cook (2005) is further complicated by retesting transfer with the same stimuli (without statistical support to rule out learning of the transfer stimuli, criterion 2), questionable novelty of some colors tested during transfer (criterion 1), and the use of two stimulus pairs presented simultaneously (i.e., 4 stimuli) in the procedure. This latter complication may be on a slightly different level, provided the pigeons were doing what people would do if they were instructed to "choose the pair with two different stimuli." Psychophysically, this would be a two-alternative forced choice task (2AFC), whereas the single pair case would be analogous to a psychophysical Yes-No task. Detectability in these two tasks is related by the square root of two, with the 2AFC task being easier than the Yes-No task (Green & Swets, 1966; Smith & Duncan, 2004). The only real potential problem with the S/D task with two pairs is whether the pigeons are doing something other than what humans would do in this task (which is difficult to know without comparative data). Because abstract-concept learning with the two-item S/D task has remained elusive over the past 30 years, researchers have used multi-item S/D discriminations that vary from the traditional two-item S/D discrimination (e.g., for a current review of this work see Cook & Wasserman, 2006). These discriminations have involved stimulus displays that usually involve more than two items (e.g., Cook, Katz, & Cavoto, 1997; Gibson, Wasserman, & Cook, 2006; Young & Wasserman, 1997). It is important to distinguish between two-item and multi-item S/D discriminations because the underlying mechanisms used for stimulus abstraction may be different (cf., Mackintosh, 2000). For a point of clarification, multi-item displays like those shown in Figure 1 often contain two unique items (e.g., U and +) but are not considered a two-item S/D discrimination because they repeat items (e.g., +) within a display, which results in a variety of potential mechanisms that may control behavior (e.g., global features that promote oddity or entropy). Consider findings from Cook et al. (1997) using visual search displays in which pigeons indicate whether a target item (e.g., the U in the left panel of Figure 1) is present (a different trial) or not (a same trial). This discrimination is based in part on an oddity mechanism (Cook 1992; Katz & Cook, 2000, 2003). For example, Cook et al. (1997) demonstrated that as the number of locations simultaneously used in the visual search displays decreased from 6 (3 x 2 array) to 4 (2 x 2 array) to 3 (1 x 3 array) the pigeons’ overall performance decreased respectively from 81.5% to 78.3% to 74.7% due to the target item becoming less odd on different trials. Pigeons were never tested with displays containing two stimuli (i.e., 1 x 1 array, the two-item discrimination) and it remains unclear whether the birds could accurately discriminate same from different displays under such conditions. (For a follow-up study consistent with this interpretation see Cook & Wasserman, 2006). Next, consider findings from Wasserman, Young, and colleagues. In their entropy-based S/D discrimination, pigeons typically indicate whether repeated items in a multi-item array are the same or different based on the number of repeated items in a display (e.g., Young & Wasserman, 1997). For example, as the number of different items decreased from 16 unique items (16D/0S; max entropy) to 16 identical items (0D/16S; minimum entropy) different responding decreased due to the decrease in entropy (see Figure 2). In follow-up studies, to explore the role of entropy in the task, the number of simultaneous locations used to present items in an array was systematically decreased (similar to that described for Cook et al., 1997) from sixteen to two (i.e., a two-item discrimination). The result of decreasing the number of items in an array is a decrease in the amount of entropy. When the number of items was reduced to two per trial, performance was equivalent to chance (Young, Wasserman, Hilfers, & Dalrymple, 1999) or pigeons’ responded same (Young, Wasserman, & Garner, 1997). These findings show the pigeon’s failure to respond different to small entropy values. In summary, in the cases discussed, two-item performance has either not been tested (Cook et al. 1997) or pigeons do not accurately discriminate displays of two items (Young, Wasserman, & Garner, 1997; Young et al., 1999). In our opinion, one needs to train and test subjects with displays containing only two items so that what has been called true or standard S/D concept learning can be tested (Mackintosh, 2000; Premack, 1983a). S/D concept learning is "true" when two items are present because it minimizes possible global perceptual features (e.g., oddity) that might confound the interpretation. Additionally, if the learned behavior is really based upon discriminating a difference between objects, then what is the rationale for it not applying to the simplest of all cases – two objects? The multi-item array experiments do not produce true S/D concept learning. Such experiments do indicate oddity and entropy-based strategies (Gibson, Wasserman, & Cook, 2006; Wasserman, Young, & Cook, 2004). But, for all the reasons just discussed, the interpretation of what the subjects are doing in the task is simplest when there are just two items and the subject judges whether they are same or different. Importance of Parametrically Varying Set-Size: Set-Size Functions Our approach to S/D concept learning has been to adhere to what we believe are the critical issues for abstract-concept learning and empirically rule out alternative explanations to abstract-concept learning (see Wright & Katz, 2006). We wanted to compare, as directly as possible, S/D concept learning of monkeys and pigeons. But since there was no clear indication from the literature as to the number of training pairs that might accomplish this goal, we made the number of training pairs a parameter of the experiment. Thus, we varied set size, the number of items used to construct the training pairs. Several studies had shown indications of a set-size effect on abstract-concept learning (e.g., Moon & Harlow, 1955; Overman & Doty, 1980; Weinstein, 1941; Wright et al., 1988), but none had studied the effects of a substantial range of set size on abstract-concept learning. We studied the effects of set size on abstract-concept learning from 8 to 128 items for capuchin monkeys (Wright, Rivera, Katz, & Bachevalier, 2003) and rhesus monkeys (Katz, Wright, & Bachevalier, 2002), and from 8 to 1024 items for pigeons (Katz & Wright, 2006). These comparative experiments were conducted with the same training pairs, testing pairs, choice responses, visual-angles of the displays, and performance criteria, providing one of the closest comparisons across species as diverse as monkeys and pigeons on S/D abstract-concept learning. In our S/D procedure the subjects were presented with two pictures and a white box (see Figure 3 and its caption for more details). If the pair of pictures were the same then a response (touch/peck) to the lower picture is correct. If the pair of pictures were different then a response to the white box was correct. Subjects were first trained with a small set size of 8 items (see Figure 4). There were 100 trials in a session (50 same and 50 different trials). After reaching the performance criterion, they were then transfer tested with novel items. The set size was then increased to 16, 32, 64, and 128 (and for pigeons 256, 512, and 1024) items with subsequent transfer tests at each set-size expansion (except at 16). Each transfer test lasted six sessions and contained 90 baseline (training stimuli) and 10 transfer trials. Transfer trials were constructed from novel items (pictures) never before seen by the pigeons (criterion 1). Each transfer item was tested once to avoid any learning effects that might occur if they had been repeated (criterion 2). The testing items were selected to be dissimilar from the training items and one another (criterion 1; many of these items can be seen in Wright & Katz, 2006).
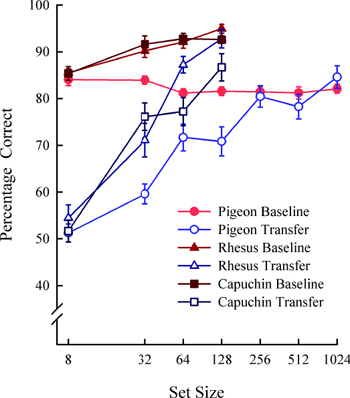
Figure 5 shows baseline and transfer performance for species across set size. Transfer performance increased with set size. These set-size functions show that monkeys fully learned (i.e., transfer performance equivalent to baseline performance) the concept by 128 items and pigeons by 256 items with displays containing only two pictures. Additionally, the animals’ transfer performance showed little or no correlation with human similarity ratings of the training and test items suggesting transfer was not a function of stimulus generalization from item features (Wright & Katz, 2006). If the subjects’ choice responses were controlled by the similarity of transfer to training stimuli then there would have been a positive correlation in as much as humans, monkeys, and pigeons perceive the stimuli in a common way.
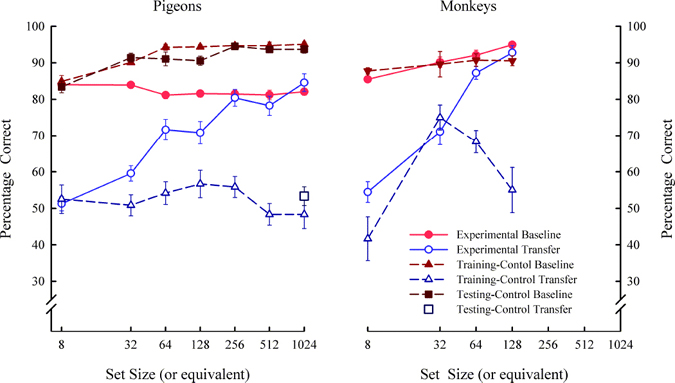
The results from control experiments corroborated these findings by ruling out training and testing experience as possible factors contributing to the increase in transfer performance. In one control experiment, we ruled out the possibility that extended training alone might produce abstract-concept learning. A training-control group of naïve pigeons and one rhesus monkey were trained with the same 8-item set as the other animals but with no set-size expansion (i.e., a fixed set of 8-items). These training-control animals were transfer tested at equivalent points in time to the set-size expansion pigeons or monkeys, so that they too would have the same experience with the same transfer pairs at the same points in training. Also, a testing-control group of pigeons were trained like the training-control group but these pigeons were tested only at the end of training (i.e., one transfer test). The testing-control animals controlled for the possibility that any transfer by training-control animals might be due to exposure of the transfer trials. The results for the experimental and control groups are shown in Figure 6. For pigeons, the control groups showed no transfer indicating that the full concept learning by the experimental group (i.e., pigeons that experienced set-size expansion) was due to set-size expansion. The absence of transfer for the training-control animals also means that the novel transfer trials themselves did not foster transfer across testing. The pigeons experienced 60 novel item pairings (90 individual pictures) across each of the 6 transfer sessions resulting in a total of 420 novel item pairings over all the transfer tests, which is somewhat of a set-size expansion only the items were not repeated. After training occurred with the fixed set of eight items, pigeons apparently needed more than one-trial training with novel stimuli and/or novel stimuli combined with training stimuli (during training) to increase their level of transfer. The same conclusion is supported by the experiments with rhesus monkeys. There is one apparent difference across these two species: the control monkey showed an increase in abstract-concept learning during the second and third test and a decrease to chance during the fourth test. The reason for this species difference is unclear, but suggests that there may be a sensitive period for developing relational learning and transferring this learning to novel stimuli (i.e., abstract-concept learning). The end result is the same for pigeons and rhesus monkeys, as overtraining with the 8-items may block relational learning. These abstract-concept learning set-size functions (Figure 5) were the first for any species. The functions help explain why previous experiments with small set sizes are likely to have resulted in claims that pigeons could not learn the abstract S/D concept. The results were also the first to show full concept learning with pigeons in a two-item S/D task. The control groups for both pigeons and rhesus monkeys were also the first of their kind to be used in a test of abstract-concept learning. Although the monkey set-size functions were of steeper slope (i.e., faster growth of concept learning) than the pigeon set-size functions, all species eventually showed full concept learning (transfer equivalent to baseline) indicating a qualitative similarity in abstract-concept learning. The somewhat slower acquisition of the S/D concept by pigeons means that they had to experience more exemplars of the rule than monkeys in order to fully transfer this rule to novel pairs of items. Why this is the case is unclear, but even humans need to be trained with several exemplars of a rule in order to fully transfer it (Chen & Mo, 2004).
Importance of Set Size, Observing Response, and Matching-To-Sample
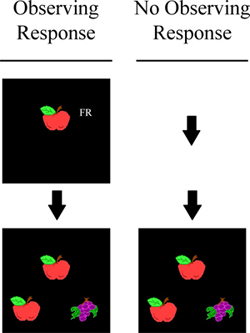
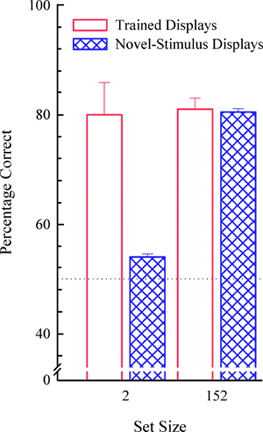
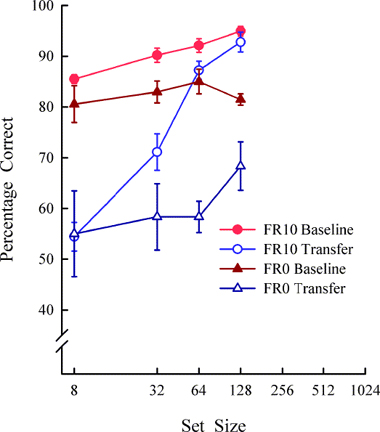
Set size is also important for pigeons in the MTS procedure (Wright et al., 1988). In typical MTS tasks, the subject is first presented a sample item (e.g., apple). After an observing response to the sample, two comparison items (e.g., apple and grapes) are typically presented (see left side of Figure 7). The correct response is to choose the comparison item (apple) that matches the sample. As with S/D, if the subject has learned the abstract matching concept (i.e., relating each comparison item to the sample item) so that it responds relationally on every trial, then it should do so on novel transfer trials and transfer performance should be as accurate as with the training stimuli. Wright et al. (1988) showed that pigeons trained with a large set size of 152 items fully learned the abstract concept and that pigeons trained with a small set size of only two items did not transfer above chance (see Figure 8). But there is another parameter, the observing response, which is important in abstract-concept learning. For example, pigeons do not need to be trained with a large set size to show full concept learning if they are required to respond to the sample item 20 times (FR20). Wright (1997, 2001) showed full transfer by pigeons with a set size of three items and an FR20. However, if the sample was pecked only once (as it was in Wright et al., 1988) or not at all (the three MTS stimuli presented all at once; see right side of Figure 7) there was no transfer. The full concept learning of the pigeons in the FR20 group from Wright (1997) was the reason we choose an FR20 for the S/D discrimination with pigeons. Clearly, there are differences between these two tasks, because the pigeons trained with 8 items and a sample response requirement of FR20 showed no abstract-concept learning. These differences between S/D and MTS could be due to any one or a combination of factors. 1) Was it a difference in the task requirements between S/D and MTS that was critical? In MTS there is always one comparison that matches the sample as in a 2AFC Signal Detection Task. 2) Does the sample observing response requirement have a different effect on MTS concept learning than on S/D concept learning? 3) Does the training set size have a different effect on MTS concept learning than on S/D concept learning? 4) Was horizontal presentation of stimuli in the MTS tasks critical to concept learning? The prior MTS experiments (Wright, 1997, 2001; Wright et al., 1988) presented stimuli from the floor of the operant chamber, as opposed to being presented vertically on the front panel. 5) Was the older EGA video monitor with its slower scan rate critical to MTS concept learning? Any or all of these factors might have played a role and deserve attention in comparing S/D and MTS procedures. In regard to our S/D procedure, the results at the very least imply that the role of the observing response for pigeons on S/D concept learning needs to be explored. Additionally, there is reason to believe we would find an observing response effect on S/D concept learning for pigeons because there was an effect with rhesus monkeys. Rhesus monkeys trained with an FR10 showed a steeper set-size function than a monkey trained with an FR0; showing an observing response effect on abstract-concept learning (see Figure 9). It would be surprising to find an effect of the observing-response requirement in rhesus monkeys, but not in pigeons. Parametrically manipulating the observing response would also be interesting for comparative reasons because in S/D abstract-concept learning pigeons were required to peck the upper item 20 times and rhesus to touch it 10 times, but capuchins did not touch the upper item at all. Capuchins were presented both items simultaneously (plus the white rectangle) and only required to make a choice response. Hence, how the observing response influences abstract-concept learning and interacts with set size across species in the S/D task is unclear. This interaction is important to understand as it may influence how the task is solved.
Discerning Different Item-Specific Strategies One of the issues addressed by Wright (1997) was how pigeons solve the MTS task when they fail transfer to novel stimuli. At the time, the explanation for failure to transfer to novel stimuli was that performance was tied to specific features of the training stimuli, i.e., item-specific learning (e.g., Carter & Werner, 1978; Wright, 1997). If subjects learn the MTS task item-specifically, then, in principle, performance can be controlled by either the configural pattern of the three stimuli or by if-then rules tied to the specific sample and comparison stimuli (e.g., Carter & Eckerman, 1975; Carter & Werner, 1978; Wright, 1997). Configural pattern learning involves learning choice responses to specific displays based on the configural gestalt or pattern (e.g., similar to an abstract painting or bed quilt) of the whole display. If-then rule learning involves learning specific stimulus-response chains to the sample and correct comparison stimulus in the MTS display, for example, "If red sample then choose red comparison", (Skinner, 1950). More than a quarter of a century ago the case was made that pigeons do not learn the MTS concept because they learn if-then rules (Carter & Werner, 1978). The possibility of configural-gestalt learning was raised by Carter & Werner (1978), but was rejected in favor of if-then rule learning with very little evidence to support either possibility.
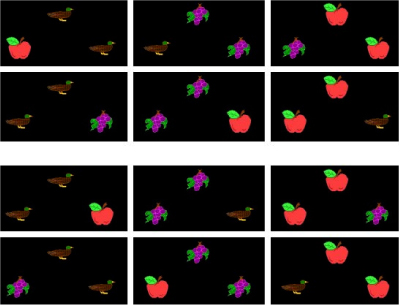
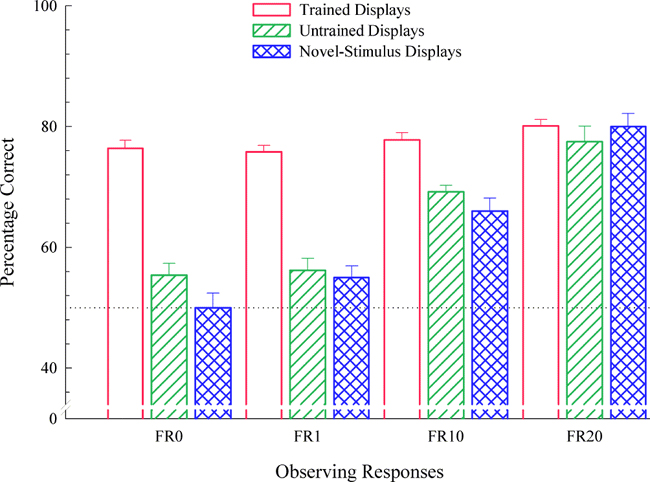
The possibility of different ways of learning has been raised off and on for many years (e.g., Carter & Werner, 1978; Lashley, 1938; Zentall & Hogan, 1974), but it was not until 1997 that it was made clear how these different possibilities might be tested (Wright, 1997). Wright devised a test for if-then rule learning. He divided the 12 possible displays constructed from three items (apple, duck, grapes) so that the roles of each item could be counterbalanced within each of two subsets of 6 displays each (see Figure 10). Using this split-set design, one set of 6 displays (e.g., the top 6 displays of Figure 10) was used in training the MTS discrimination and the other set (e.g., the bottom 6 displays of Figure 10) was used specifically to test for if-then rule learning (but were counterbalanced in the experiment). If the pigeons learned the task by if-then rules, they should have transferred their performance to the set of 6 displays not used in training, i.e., novel displays constructed from the same familiar items used in training, because the same if-then rules (e.g., "If duck then choose duck comparison") would be equally effective with either subset. If the pigeons learned the task configurally, they should have failed to transfer to the untrained displays. Wright also varied the number of observing responses (FR 0, 1, 10, or 20) to the sample across groups. The results are shown in Figure 11. When pigeons did not learn the abstract concept, they fully learned the MTS task configurally (FR0 and FR1). Pigeons that were required to peck the sample 20 times fully learned the matching concept, even with a small training set of 3 items, as shown by transfer to novel stimuli being as good as training-trial performance. These results clearly show that pigeons are capable of learning a variety of strategies and that parametric manipulation can influence what strategy is learned.
Conclusions In closing, we would like to make a few comments about the importance of tasks which can be solved by different strategies. Such tasks are important to study because they can help reveal the cognitive flexibility of human and nonhuman animals. MTS and S/D are two such tasks that can be solved by either relational or item-specific strategies (e.g., Carter & Warner, 1978; Cumming & Berryman, 1965; Wright, 1997). A variety of species including capuchin monkeys (Wright et al., 2003), chimpanzees (Oden, Thompson, & Premack, 1988), dolphins (Herman, Hovancik, Gory, & Bradshaw, 1989), humans (Weinstein, 1941), parrots (Pepperberg, 1987), rhesus monkeys (Wright et al., 1984), sea lions (Kastak & Schusterman, 1994), and even pigeons (Katz & Wright, 2006; Wright, 1997) can solve the MTS and/or S/D task either item-specifically or relationally. The pervasiveness of such multiple strategy use is clearly seen across the human life-span in a wide variety of tasks including arithmetic, time telling, serial recall, spelling, and conservation (for a review see Siegler, 1996). For example, when solving addition problems children (5-7 years old) can use a combination of the MIN (counting up by ones from the larger addend by the smaller addend), decomposition (transforming a difficult problem into two simpler ones), guessing, or retrieval (accessing the answer from memory) strategies. Hence, how animals learn the MTS and S/D tasks may share similar processes compared with how humans generally learn and solve problems. Understanding how these processes are quantitatively and qualitatively same or different across species can advance our knowledge about how animals process and think about things. Such progress may decrease the commonly misperceived gap between human higher-order cognitive processing and that of nonhuman animals. References Bitterman, M. E. (1965). Phyletic differences in learning. American Psychologist, 20, 396-410. Bitterman, M. E. (1975). The comparative analysis of learning. Science, 188, 699-709. Blaisdell, A. P., & Cook, R. G. (2005). Two-item same-different concept learning in pigeons. Learning & Behavior, 33, 67-77. Bovet, D., & Vauclair, J. (2001). Judgment of conceptual identity in monkeys. Psychonomic Bulletin & Review, 8, 470–475. Carter, D. E. & Eckerman, D. A. (1975). Symbolic matching by pigeons: Rate of learning complex discriminations predicted from simple discriminations. Science, 187, 662-664. Carter, D. E., & Werner, J. T. (1978). Complex learning and information processing in pigeons: A critical analysis. Journal of the Experimental Analysis of Behavior, 29, 565-601. Chen, Z., & Mo, L. (2004). Schema induction in problem solving: A multidimensional analysis. Journal of Experimental Psychology: Learning, Memory, and Cognition, 30, 583-600. Cook, R. G. (1992). The acquisition and transfer of texture visual discriminations by pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 18, 341-353. Cook, R. G. (2002). Same-different concept formation in pigeons. In M. Bekoff, C. Allen, & G. M. Burghardt (Eds.), The cognitive animal (pp. 229-237). Cambridge, MA: MIT. Cook, R. G., Katz, J. S., & Cavoto, B. R. (1997). Pigeon same different concept learning with multiple stimulus classes. Journal of Experimental Psychology: Animal Behavior Processes, 23, 417-433. Cook, R. G., Kelly, D. M. & Katz, J. S. (2003). Successive two-item same-different discrimination and concept learning by pigeons. Behavioral Processes, 62, 125-144. Cook, R. G. & Wasserman, E. A. (2006). Relational discrimination learning in pigeons. In E. A. Wasserman & T. R. Zentall (Eds.), Comparative cognition (pp 307-324). New York, NY: Oxford University Press. Cumming, W. W., & Berryman, R. (1965). The complex discriminated operant: Studies of matching-to-sample and related problems. In D. I. Mostofsky (Ed.), Stimulus generalization (pp. 284-330). Stanford, CA: Stanford University Press. D’Amato, M. R., Salmon, D. P., & Colombo, M. (1985). Extent and limits of the matching concept in monkeys (Cebus apella). Journal of Experimental Psychology: Animal Behavior Processes, 11, 35-51. Daehler, M. W., & Bukatko, D. (1985). Cognitive development. New York: Alfred A. Knopf. Gibson, B. M., Wasserman, E. A., & Cook, R. G. (2006). Not all same-different discriminations are created equal: Evidence contrary to a unidimensional account of same-different learning. Learning and Motivation, 37, 189-208. Green, D. M. & Swets, J. A. (1966). Signal detection theory and psychophysics. New York: Wiley. Herman, L. M., Hovancik, J. R., Gory, J. D., & Bradshaw, G. L. (1989). Generalization of visual matching by a Bottlenosed Dolphin (Tursiops truncates): Evidence for invariance of cognitive performance with visual and auditory materials. Journal of Experimental Psychology: Animal Behavior Processes, 15, 124-136. Herrnstein, R. J. (1990). Levels of stimulus control: A functional approach. Cognition, 37, 133-166. Herrnstein, R. J., Loveland, D. H., Cable, C. (1976). Natural concepts in pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 2, 285-302. James, W. (1950). The principles of psychology (Vol. 1). New York: Dover Publications. (Original work published 1890) Kamil, A. C. (1988). A synthetic approach to the study of animal intelligence. In D. W. Leger (Ed.), Nebraska Symposium on Motivation, 35, 257-308. Kastak, D. & Schusterman, R. J. (1994). Transfer of visual identity matching-to-sample in two California sea lions (Zalophus californians). Animal Learning & Behavior, 22, 427-435. Kastak, C. R. & Schusterman, R. J. (2002). Sea lions and equivalence: Expanding classes by exclusion. Journal of the Experimental Analysis of Behavior, 78, 449-465. Katz, J. S., & Cook, R. G. (2000). Stimulus repetition effects on texture-based visual search by pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 26, 220-236. Katz, J. S., & Cook, R. G. (2003). The multiplicity of visual search strategies in pigeons. In Sal Soraci, Jr. and Kimiyo Murata-Soraci (Eds.), Visual information processing. Westport, CT: Greenwood Publishing Group. Katz, J. S., & Wright, A. A. (2006). Mechanisms of same/different abstract-concept learning by pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 32, 80-86. Katz, J. S., Wright, A. A., & Bachevalier, J. (2002). Mechanisms of same/different abstract-concept learning by rhesus monkeys (Macaca mulatta). Journal of Experimental Psychology: Animal Behavior Processes, 28, 358–368. Lashley, K. S. (1938) Conditional reactions in the rat. Journal of Psychology, 6, 311-324. Mackintosh, N. J. (1988). Approaches to the study of animal intelligence. British. Journal of Psychology, 79, 509-525. Mackintosh, N. J. (2000). Abstraction and discrimination. In C. Heyes & L. Huber (Eds.), The evolution of cognition (pp. 123-141). Cambridge, MA: MIT. Macphail, E. M. (1985). Vertebrate intelligence: The null hypothesis. Philosophical Transaction of the Royal Society of London, Series B, 308, 37-51. Marcus, G. F., Vijayan, S., Bandi Rao, S., & Vishton, P. S. (1999). Rule learning by seven-month-old infants. Science, 283, 77-80. Medin, D. L., (1989). Concepts and conceptual structure. American Psychologist, 44, 1469-1481. Moon, L. E., & Harlow, H. F. (1955). Analysis of oddity learning by rhesus monkeys. Journal of Comparative and Physiological Psychology, 48, 188-195. Oden, D. L., Thompson, R. K. R., & Premack, D. (1988). Spontaneous transfer of matching by infant chimpanzees (Pan troglodytes). Journal of Experimental Psychology: Animal Behavior Processes, 14, 140-145. Overman, W. H., Jr., & Doty, R. W. (1980). Prolonged visual memory in macaques and man. Neuroscience, 5, 1825-1831. Papini, M. R. (2002). Pattern and process in the evolution of learning. Psychological Review, 109, 186–201. Pepperberg, I. M. (1987). Acquisition of the same/different concept by an African Grey parrot (Psittacus erithacus): Learning with respect to categories of color, shape, and material. Animal Learning and Behavior, 15, 423-432. Piaget, J., & Inhelder, B. (1969). The psychology of the child (H. Weaver, Trans.). New York: Basic Books. (Original work published 1966). Premack, D. (1978). On the abstractness of human concepts: Why it would be difficult to talk to a pigeon. In S. H. Hulse, H. Fowler, & W. K. Honig (Eds.), Cognitive processes in animal behavior (pp. 423-451). Hillsdale, NJ: Erlbuam. Premack, D. (1983a). Animal cognition. Annual Review of Psychology, 34, 351-362. Premack, D. (1983b). The codes of man and beasts. The Behavioral and Brain Sciences, 6, 125–167. Santiago, H. C., & Wright, A. A. (1984). Pigeon memory: Same/different concept learning, serial probe recognition acquisition, and probe delay effects on the serial-position function. Journal of Experimental Psychology: Animal Behavior Processes, 10, 498-512. Shettleworth, S. J. (1998). Cognition evolution and Behavior. New York: Oxford University Press. Siegler, R. S. (1996). Emerging minds: The process of change in children’s thinking. New York: Oxford University Press. Skinner, B. F. (1950). Are theories of learning necessary? Psychological Review, 57, 193-216. Smith, D. G., & Duncan, M. J. J. (2004). Testing theories of recognition memory by predicting performance across paradigms. Journal of Experimental Psychology: Learning, Memory, & Cognition, 30, 615-625. Thomas, R. K. (1980). Evolution of intelligence: An approach to its assessment. Brain, Behavior and Evolution, 17, 454–472. Thomas, R. K. (1996). Investigating cognitive abilities in animals: unrealized potential. Cognitive Brain Research, 3, 157-166. Thompson, R. K. R. (1995). Natural and relational concepts in animals. In H. L. Roitblat and J.-A. Meyer (Eds.) Comparative approaches to cognitive science (pp. 175-224). Cambridge, MA: MIT Press. Thompson, R. K. R., & Oden, D. L. (2000). Categorical perception and conceptual judgments by nonhuman primates: the paleological monkey and the analogical ape. Cognitive Science, 24, 363–396. Thompson, R. K. R., Oden, D. L., & Boysen, S. T. (1997). Language-naive chimpanzees (Pan troglodytes) judge relations between relations in a conceptual matching-to-sample task. Journal of Experimental Psychology: Animal Behavior Processes, 23, 31-43. Thorndike, E. L. (1998). Animal intelligence. Experimental studies. Thoemmes Press, Bristol, UK (Original work published in 1911). Tooby, J., & Cosmides, L., 2000. Toward mapping the evolved functional organization of mind and brain. In M.S. Gazzaniga (Ed), The new cognitive neurosciences: Second Edition (pp. 1167-1178). Cambridge, MA: MIT Press. Urcuioli, P. J. (2006). Responses and acquired equivalence classes. In E. A. Wasserman & T. R. Zentall (Eds.), Comparative Cognition (pp 405-421). New York, NY: Oxford University Press. Vaughn, W., Jr. (1988). Formation of equivalence sets in pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 14, 36-42. Wasserman, E. A., DeVolder, C. L., & Coppage, D. J. (1992). Non-similarity-based conceptualization in pigeons via secondary or mediated generalization. Psychological Science, 3, 374-379. Wasserman, E. A., Kiedinger, R. E., Bhatt, R. S. (1988). Conceptual behavior in pigeons: Categories, subcategories, and pseudocategories. Journal of Experimental Psychology: Animal Behavior Processes, 14, 235-246. Wasserman, E. A., Young, M. E., & Cook, R. G. (2004). Variability discrimination in humans and animals. American Psychologist, 59, 879-890. Weinstein, B. (1941). Matching-from-sample by rhesus monkeys and by children. Journal of Comparative Psychology, 31, 195-213. Wright, A. A. (1991). Concept learning by monkeys and pigeons. In M. Corballis, K. G. White, and W. Abraham (Eds.), Memory mechanisms: A tribute to G. V. Goddard (pp. 247-273). Hillsdale, N. J.: Erlbaum. Wright, A. A. (1997). Concept learning and learning strategies. Psychological Science, 8, 119-123. Wright, A. A. (2001). Learning strategies in matching to sample. In. R. G. Cook (Ed.), Avian visual cognition. [On-line]. Available: www.pigeon.psy.tufts.edu/avc/. Wright, A. A., Cook, R. G., Rivera, J. J., Sands, S. F., & Delius, J. D. (1988). Concept learning by pigeons: Matching-to-sample with trial-unique video picture stimuli. Animal Learning & Behavior, 16, 436-444. Wright, A. A., & Delius, J. D. (2005). Learning processes in matching and oddity: The oddity preference effect and sample reinforcement. Journal of Experimental Psychology: Animal Behavior Processes, 31, 425-432. Wright, A. A., & Katz, J. S. (2006). Mechanisms of same/different concept learning in primates and avians. Behavioural Processes, 72, 234-254. Wright, A. A., Rivera, J. J., Katz, J. S., & Bachevalier, J. (2003). Abstract-concept learning and list-memory processing by capuchin and rhesus monkeys. Journal of Experimental Psychology: Animal Behavior Processes, 29, 184-198. Wright, A. A., Santiago, H. C., Urcuioli, P. J. and Sands, S. F. (1984). Monkey and pigeon acquisition of same/different concept using pictorial stimuli. In M. L. Commons and R. J. Herrnstein (Eds.), Quantitative analysis of behavior: Vol. IV. Discrimination processes (pp. 295-317). Cambridge, Ma: Ballinger. Young, M. E., & Wasserman, E. A. (1997). Entropy detection by pigeons: Response to mixed visual displays after same-different discrimination training. Journal of Experimental Psychology: Animal Behavior Processes, 23, 157-170. Young, M. E., Wasserman, E. A., & Garner, K. L. (1997). Effects of number of items on the pigeon’s discrimination of same from different visual displays. Journal of Experimental Psychology: Animal Behavior Processes, 23, 491-501. Young, M. E., Wasserman, E. A., Hilfers, M. A., & Dalrymple, R. M. (1999). The pigeon’s variability discrimination with lists of successively presented visual stimuli. Journal of Experimental Psychology: Animal Behavior Processes, 25, 475-490. Zentall, T. R., & Hogan, D. E. (1974). Abstract concept learning in the pigeon. Journal of Experimental Psychology, 102, 393-398. |


Jeffrey S. Katz and Kent D. Bodily, Department of Psychology, Auburn University; Anthony A. Wright, University of Texas Health Science Center at Houston, Houston, Texas.
|