| <<Vol. 2 Table of Contents | |
|
|
|
|
2007 |
Volume 2, pp 67-78 |

Individual Differences and Animal Personality
Charles Locurto
College of the Holy Cross
|
|
|
As illustrated in the above quote by Yerkes it is commonplace to regard individual animals as possessing distinct personalities. Yerkes went on to say in that same article, "One may as readily identify a familiar ape among many by its personality as mirrored in behavior as by its physical appearance" (1939, p. 111). I am guessing that anyone with extensive contact with almost any species, certainly with any primate species, would say something similar. As a consequence of that widespread opinion, while personality may not be a topic that presently attracts much attention within comparative cognition, the study of personality in nonhumans is in fact abundant.
Two aspects of this review of animal personality studies deserve mention at the outset. The first is a definitional point: Personality is often distinguished from temperament. The study of temperament originated in observations of children in which consistent behavioral styles could often be identified at an early age (Clarke & Boinski, 1995). As a result, temperament is usually regarded as consisting of fundamental behavioral dispositions (e.g., impulsive, shy, curious) that are unlearned (Box, 1999; Kagan, Saidman, & Arcus, 1992). The term personality is sometimes reserved for dimensions of behavior that are more socially influenced and need to be observed in a variety of situations before being labeled accurately (e. g., neurotic, conscientious, confident). At times it is difficult to distinguish between personality and temperament. In animal studies the terms appear at times to be used interchangeably (Jones & Gosling, 2005). In this review the term personality will be used to encompass both of these behavioral styles. Also, this review of animal studies of personality will concentrate on primates due to the vastly more developed literature in this biological order compared to others. It should be said however that attributions of personality have been made to a number of nonprimate species including dogs (Canis familiaris; Jones & Gosling, 2005), birds (Zebra finch (Poephilia guttata); Figueredo, Cox, & Rhine, 1995), and even squid (Euprymna tasmanica; Sinn & Moltschaniwskyj, 2005) and the octopus (Octopus rubescens; Mather & Anderson, 1993). Taken in sum, studies of personality in nonhuman animals constitute an impressively broad and developed literature.
Dogs might be accorded a privileged status for an audience interested in comparative cognition inasmuch as it was Pavlov (1906; 1941) who laid a foundation for the study of associative processes and also developed what was arguably the first systematic typology for personality in a nonhuman species. Pavlov identified 4 types that he considered to be exemplars from a larger set of personality profiles. The exemplar types are presented in Table 1. Pavlov’s typology derived from three properties of the nervous system: Force, Equilibrium, and Mobility, each of which is briefly defined in the Table. The resulting mixture of these three properties resulted in the exemplar types. Interestingly, this system bears similarities to the ancient Greek system of personality types based upon the four humours: choleric (Excitable), sanguine (Lively), phlegmatic (Quiet), and Melancholic (Inhibited).
Table 1: Pavlov’s Typology
|
||||||||||||||||
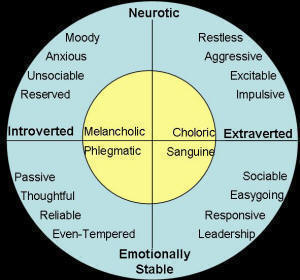
Pavlov’s work on personality types in dogs has encouraged a considerable amount of additional work on this topic. In a recent review Jones & Gosling (2005) noted that in the period 1934 – 2004, 51 empirical studies of personality in dogs had been published. Pavlov’s typology was also influential in the later development of Eysenck’s (1967) personality theory, which in its original form captured different personality types within an orthogonal two dimension system consisting of neuroticism/emotional stability on one dimension and extraversion/introversion on the other, as illustrated in Figure 1. Eysenck later added a third dimension, psychoticism, after extensive observations of human psychiatric patients.
 |
|
Figure 1. Illustration of Eysenck’s two dimensional personality typology. (Redrawn from Eysenck & Eysenck, 1958). |
Pavlov was clear that these personality types were markers for different styles of associative learning (Gray, 1964). The Excitable type, for example, exhibited strong excitatory conditioning but a limited ability to develop inhibitory connections. As a consequence, excitatory learning was rapid in these animals but learning that required a pronounced interplay between excitatory and inhibitory processes was often slow. The Lively type was the most well-balanced of the four types with respect to the interplay and transition between excitation and inhibition. As a consequence these animals evidenced rapid associative learning. Pavlov’s Quiet type exhibited consistent but slow learning, the result of low mobility between excitation and inhibition. The weak type, the only type in which inhibition dominated excitation, as might be expected, exhibited slow and difficult excitatory conditioning. These animals possessed little tolerance for intense stimulation and were easily thrown into what Pavlov termed protective inhibition to prevent cortical damage.
Pavlov’s work on personality types in dogs was extensive but it was by no means singular in identifying types of personality in a nonhuman species. As will be seen, it is by now settled that there are indeed identifiable behavioral types, and that these types often correspond to comparable types that have emerged from studies of human personality. Additionally, a number of neurobiological correlates of personality types have been identified in animals that appear to correspond to processes identified in humans. A question of special importance to students of comparative cognition is whether an understanding of these types and their neurobiological correlates is at all relevant to understanding cognition. For Pavlov the answer clearly was yes, but admittedly there has been relatively little work that identifies correspondences between personality differences and differences in cognition. An answer to this question is difficult to come by, but I will make the case that the answer is yes.
Descriptions of Personality Types in Nonhuman Primates
Personality may be difficult to define in the abstract but any definition assumes that individual differences demonstrate stability across varying contexts. Just as we would be more likely to label someone intelligent if they demonstrated that quality in several situations, we would tend to label someone extraverted if they appeared that way in more than one circumstance. This idea has led to the methodological strategy of using a battery of measures in personality assessments that can provide multiple markers to characterize the personality of an individual. In the case of animal studies these batteries are typically in the form of behavioral codings or subjective ratings of traits (Gosling, 2001). Data analysis often proceeds by some form of factor or principal components analysis that identifies clusters of traits or behaviors that covary across individuals. It should be added that personality is regarded principally as composed of processes that are distinct. As a consequence models of personality tend not to be hierarchical with a single organizing factor at the apex as is common in studies of human cognitive differences, but are instead composed of factors that are essentially orthogonal to each other. Eysenck’s model (Eysenck & Eysenck, 1958) illustrates this characteristic as does the more recent Five Factor Model that posits dimensions of Neuroticism, Extraversion, Agreeableness, Conscientiousness, and Openness to Experience (Goldberg, 1990).
The first requirement of any attempt to assess personality is that the measures be reliable, that is, replicable across different raters or within the same rater in a test-retest design. In studies of human personality reliabilities tend to be in the range .50 - .85, with most reliabilities falling between .70 - .85 (Akien, 1988). A recent review of animal personality studies by Gosling (2001) indicated that reliabilities fell close to the average in studies of human personality. Test-retest reliabilities averaged .73, a value that is within the range of values observed in studies of human personality. Median weighted inter-observer reliabilities averaged .52 across 375 separate reliability estimates, a value that again compares well with the average interobserver reliability reported in studies of human personality. Interobserver correlations represent agreement among judgments of the same construct. As a consequence they need not be squared, as in computing a coefficient of determination, but are directly interpreted as indicating shared variance (Ozer, 1985).
 |
|
Figure 2. Rhesus macaque. Image retrieved August 10, 2006, from www.pin.primate.wisc.edu/fs/sheets/images/~ron/index.html. Copyright 2006, Franz de Waal. Reprinted with permission. |
A second requirement that is more difficult to assess is validity, the extent to which a given measure is tapping the construct that it is assumed to be marking. How can we know that avoidance of a novel object placed in the home cage, a common experimental method for assessing an aspect of neuroticism in animals, is indeed marking neuroticism? To be sure, meeting this requirement is also problematic in studies of human personality, inasmuch as it is difficult to establish the benchmark measure of a given construct, that is, the measure against which the validity of other measures can be judged. In the absence of benchmark measures, either for humans or animals, an alternate strategy is a form of construct validity which examines the correlations, the convergences, among several measures that presumptively tap the same construct. In the case of animal studies this type of validity might take the form of correlations between several rated dimensions each of which is thought to tap aspects of neuroticism, or it might involve correlations between experimental manipulations, behavioral codings, and subjective ratings of different aspects of neuroticism. As an example of this strategy, it has been demonstrated in rhesus macaques (Macaca mulatta, see Figure 2) that personality ratings correlated substantially with codings of social behaviors taken in contexts different from those in which the ratings took place, and that these consistencies were evident for several years following the initial ratings (Capitano, 1999). While there is less information on validity than on reliability, and while validity correlations are generally lower than reliability measures, the available data suggest that correlations between different measures of the same construct are reasonably high, in the range of .30 to .50. This range is comparable to validity measures observed in studies of human personality (e.g., Gosling, 2001; Stevenson-Hinde & Zung, 1978).
The strength of reliability and validity measures reported in studies of animal personality supports Pavlov’s intuitions about the presence of systematic types or dimensions. That said, it should also be said that the animal literature is less coherent than is the human literature with respect to the confirmation of similar personality dimensions across different studies, either within or between species, although there is evidence of replications of some factor structures within species (see Capitano & Widaman, 2005, in rhesus macaques). Interestingly, there are factors that do not appear to be symmetrically represented in animals and humans. Conscientiousness, a factor that consistently sorts out in human studies, has been observed in chimpanzees but not in other primates including Orangutans (Pongo pygmaeus and Pongo abelii; Weiss, King, & Perkins, 2006). This factor is marked in chimps by attention, goal-directedness, and organized/disorganized behavioral patterns, making it in all likelihood more narrowly defined in chimps than in humans. Dominance often appears in primate studies and, as will be discussed, has been the subject of a considerable amount of work. It is seldom extracted as a separate factor in human studies although some aspects of extraversion may mark this factor. As might be expected this factor is usually related to an individual’s rank in a colony hierarchy (e.g., King & Figeuredo, 1997). There are also factors that might be thought of as uniquely human but may have nonhuman counterpart. Psychopathy is a factor of this sort. It is typically marked by traits such as dishonesty, superficial charm, and guiltlessness. There is work with chimpanzees (Pan troglodytes) suggesting that some markers of this trait such as poor impulse control, bluff displays, and failure to form intimate attachments, can be reliably observed (Lilienfeld et al., 1999).
 |
|
Figure 3. Vervet monkeys. Image retrieved August 18, 2006, from www.biochem.ucl.ac.uk/~shepherd/vervet.html. Copyright, 2006, Adrian Shepherd. Reprinted with permission. |
Conscientiousness and dominance may not be symmetrically represented in human and nonhuman primates but it is common in studies of nonhuman primates to find factors that appear to correspond to those that compose the modern Five Factor Model (Itoh, 2002). Extraversion, for example, often emerges in studies of both humans and animals (in Orangutans; Weiss et al., 2006). In animals this dimension is often marked by traits such as exploration and energy. Agreeableness or sociability is also often extracted, with markers such as affability and affection on the positive end of this factor, and aggression on the negative end (e.g., in lion-tailed macaques (Macaca silenus; Roeff, Susman, & Strube, 2005). Neuroticism is also observed in a variety of primate species with behavioral markers such as impulsivity, fearfulness, and emotional reactivity (Gosling, 2001). Social impulsivity, in particular, has often been observed using a form of the intruder paradigm. Interestingly, in vervet monkeys (Cercopithecus aethiops sabaeus; see Figure 3) it has been found that the highest ranking males were more likely to score in the moderate range in their approach to an intruder introduced along the periphery of their home enclosure, whereas low ranking males were more likely to register more extreme scores in either direction (Fairbanks, 2001). This outcome suggests a nuanced view of how aspects of personality map on to social ranking, a point to which I will return.
Characteristics of neuroticism, particularly those features that involve impulsivity or reactivity to novel stimuli and/or conspecifics, constitute some of the most well-studied aspects of primate personality. An extensive series of studies by Suomi and others has documented in rhesus macaques that these behavioral styles emerge early in life and are highly heritable (Higley & Suomi, 1989; Suomi, 1991, 1997). The most prominent style appears in a subset of approximately 20 percent of rhesus macaques that are labeled "reactive" in that their responses to changes in their physical and social environment are extreme, both behaviorally and physiologically. These animals are less exploratory, more socially inhibited, and more anxious than other members of their colony. They can be identified early in life as infants that are reluctant to leave their mothers. It has been observed that this behavioral style may provide a compelling model for human reactive depression in that both reactive rhesus macaques and reactive human depressives respond strongly to environmental challenges, evidence high anxiety and fear in novel situations, and develop depressive reactions in response to prolonged exposure to stress (Suomi, 1997).
An additional 5 to 10 percent of rhesus macaques can be described as impulsive and risk takers, especially in situations that commonly involve play or social aggression (Box, 1999; Capitano & Widaman, 2005; Fairbanks, 2001). These individuals appear less socially oriented than their age mates and seem to exhibit less behavioral control. They migrate out of their home colony earlier than their age mates, and they are more susceptible to injuries from falls and aggressive encounters. Not surprisingly, they have relatively high mortality rates.
The remainder of a rhesus macaque colony consists of individuals that are less reactive, less impulsive, and more appropriately exploratory than these two extreme groups. It has been speculated that these more emotionally stable animals enjoy several advantages over members of the reactive and impulsive subgroups. They tend to establish more stable and supportive social networks than their extreme counterparts. They have more opportunities to interact successfully with adults, and as a consequence they are more likely to be afforded protection in times of threats from predation. They also tend to gain access to better food patches. On this point, there is evidence that these animals develop more successful, energetic foraging strategies than reactive or impulsive animals (Box, 1999).
It should not escape attention that these personality types in rhesus macaques bear some similarities to Pavlov’s original typology. The subset of impulsive, risk taking monkeys appear to have much in common with Pavlov’s Excitable type, although Pavlov’s types did not include criteria derived from social contexts, but were instead developed to correspond to different manners of associative learning. The reactive style appears to be similar to Pavlov’s Inhibited type, although again in primates this type is derived from evidence that includes responsiveness in social situations – intruder threats, peer interactions, and mother-child exchanges. It is not clear presently whether Pavlov’s remaining two types, the Lively and the Quiet types, find their counterpart in primate studies. The large percentage of individuals in primate colonies that are characterized as emotionally balanced and not belonging to one of the extreme groups, perhaps upwards of two-thirds of a colony, have not been further delineated. It would not be surprising if additional work on this group identifies some behavioral styles that are commensurate with Pavlov’s idea that these more emotionally balanced individuals can be further subdivided.
The neurobiology of personality
As expected, there are differences in the physiological substrates of these different behavioral styles. Highly impulsive monkeys evidence chronically low central serotonin metabolism (Suomi, 1997). The serotonergic system has been implicated in numerous aspects of brain functioning, ranging from the regulation of food intake and circadian rhythms to the expressions of mood, including anxiety and aggression (Reif & Lesch, 2003). It has been shown that this low serotonin profile in monkeys resembles Type II alcoholism in humans in terms of covarying with excessive alcohol consumption, social alienation, and with a type of aggression that is violent and unrestrained (Higley & Bennett , 1999). In humans, serotonin dysfunction has also been associated with borderline personality disorder and suicide (Reif & Lesch, 2003). Interestingly, in the nonhuman primate species that have been studied approximately 10 – 20 percent of individuals will consume alcohol at high rates. This percentage is likely larger than the subgroup of impulsive individuals, suggesting that behavioral styles other then impulsivity may be associated with susceptibility to alcoholism. This percentage is quite similar to the percent of humans who abuse alcohol during some period of their lives (Higley & Bennett, 1999).
The most studied biological correlates of different personalities in nonhuman primates concern those that attend exposure to stress. A standard method for inducing stress in primates is through social separation (e.g., Suomi, 1997). When rhesus macaques are stressed in this manner, clear differences emerge between reactive individuals and their more evenly tempered counterparts. Under stress, reactive monkeys evidence more dramatic changes in sympathetic nervous system functioning and hypothalamic-pituitary-adrenal (HPA) activity as assessed principally by levels of adrenocorticotropic hormone (ACTH) and cortisol. ACTH stimulates the adrenal cortex thereby influencing secretion of glucocorticoids such as cortisol. Cortisol increases blood pressure, blood sugar levels, and suppresses the immune system. For days following a social separation test the sleep and cardiac patterns remain altered in reactive monkeys. These altered patterns can be reversed by treatment with antidepressants. Work with tufted capuchin (Cebus apella) juveniles has shown that elevated levels of cortisol following a stressful stimulus are associated with lower levels of play and home-cage exploration, and with higher levels of proximity with mothers. Additionally, high cortisol reactivity correlated positively with personality traits such as fearfulness, insecurity, and submissiveness, and correlated negatively with traits such as confidence, curiousity, and effectiveness (Byrne & Suomi, 2002).
 |
|
Figure 4. Sapolsky and friend, an olive baboon. Copyright, 2006, Robert Sapolsky. Reprinted with permission. |
Other work with reactive rhesus macaques indicates that these animals also evidence extreme asymmetric right frontal electrical activation in response to stress. This pattern is also found in adult humans who exhibit negative affect and anxious dispositions (Kalin, 2003). This asymmetrical pattern is also found in children diagnosed as behaviorally inhibited. These children are characterized as avoiding unfamiliar events and people. In the face of novel challenges they tend to seek the protection of caregivers. This shy/highly-introverted pattern evidences some level of continuity throughout childhood, and to some extent is predictive of adult personality (Fox et al., 2005). It has been suggested that frontal asymmetries of this sort can be understood motivationally, as marking differences in the balance between appetitive/approach and withdrawal/avoidance activation. Left frontal activation predicts approach tendencies whereas right frontal activation is more likely associated with avoidance (Fox et al., 2005).
Basal cortisol levels have also been related to the dominance hierarchy in primates. Perhaps the most influential work in this regard has been done by Sapolsky in an extensive study of the social hierarchy in olive baboons (Papio anubis; Ray & Sapolsky, 1992; Sapolsky & Ray, 1989; Sapolsky, 1990; see Figure 4). The original conclusion about the relationship between biological profile and dominance was that dominant males in stable hierarchies had lower basal concentrations of cortisol. Later work revealed that this profile was found only in males with a certain style of dominance, one characterized by a relatively deliberate, measured response to environmental challenges. These males had the ability to distinguish between threatening and more neutral interactions with male rivals, and were better able to initiate successful fights when necessary. They were also skilled at displacing aggression, finding a safe outlet for their aggressive arousal rather than confronting other high-ranking males directly.
There was a second style of dominance evident in these baboon groups. This second subset of males lacked the well-developed social skills of the first group. Members of this group were instead highly responsive to environmental challenges, more along the lines of the impulsive type of rhesus macaques identified by Suomi (1997). Interestingly, this subgroup of males did not necessarily engage in more fighting. Cortisol levels did not correlate with the overall frequency of male-male aggression. Males who initiated the highest percentage of their fights had, in fact, lower basal concentrations of cortisol than low-initiating males. For this subset of highly responsive dominant males basal cortisol levels were as high as those found typically in subordinate males. In subordinates high basal cortisol concentrations tended to covary with suppression of high-density lipoproteins (HDL), the so-called "good" cholesterol that is associated with a lowered risk of coronary artery disease in humans. High cortisol levels also covaried with steroid diabetes, hypertension, and immunosuppression in these subordinates.
As Sapolsky and Ray (1989) noted, the behavioral style of the dominant males with low-cortisol profiles would tend to mitigate the impacts of stress and as a consequence might allow them more control over potentially stressful situations. Perhaps not surprisingly, males with this pattern of dominance lasted longer in their positions of high rank than did their more highly-responsive counterparts. This outcome dovetails with a considerable body of research indicating that is not stress per se but often the unpredictability of stress that produces psychopathological consequences (Seligman & Weiss, 1980). Interestingly, the advantages owing to this low-cortisol profile were absent during periods of group instability. During these periods, formerly low-cortisol males evidenced basal cortisol levels as high as subordinates.
It should be added that the social rank of individual baboons in Sapolsky’s studies changed over time, although the physiological profile of these animals often remained relatively constant. Moreover, cortisol measures taken prior to the formation of social groups were at best moderate predictors of eventual rank (Sapolsky, 1990). These patterns indicate that while the physiological profiles identified by Sapolsky tend to covary with social rank, they are not unique predictors of it. Complicating this picture further is that Ray and Sapolsky (1992) identified three distinctive behavioral styles, each of which was associated with low cortisol levels: a sexually-engaged style that included high rates of grooming and copulation, a social affiliative style that was marked in part by interactions with non-estrus females and infants, and a third style that was marked by the ability to judge threatening from nonthreatening interactions with rivals.
There are other physiological correlates of social rank. In humans cortisol levels are often evaluated against concurrent serotonin levels given that imbalances between the two are often associated with negative behavioral outcomes like depression and sleep disorders (Rang, Dale, Ritter, & Moore, 2003). As might be inferred from the prior discussion of the outcomes of elevated cortisol levels, low serotonin levels in rhesus macaques are associated with the same behavioral style that is marked by high cortisol levels in terms of impulsivity, low levels of grooming and other social activities, and violent aggression (Dingemanse & Réale, 2005). Serotonin received considerable attention in studies of animal dominance following the demonstration that in vervets, males became dominant in their group following treatment with the amino acid precursor of serotonin, tryptophan, and assumed subordinate roles when treated with fenfluramine, a serotonin depleting agent (Raleigh, McGuire, Brammer, Pollack & Yuwiler, 1991).
In Cynomolgus monkeys (Macaca fascicularis) it has been shown that levels of homovanillic acid (HVA), the principal dopamine metabolite in the primate central nervous system, correlated positively with ratings of dominance in both males and females (Kaplan, Manuck, Babette, & Mann, 2001). The involvement of dopamine is not as yet well understood but its relationship to dominance is interesting. Dopamine has traditionally been associated with the brain’s pleasure systems and is released in conjunction with activities such as feeding, sex, and novelty seeking, including the intake of certain addictive drugs like cocaine or amphetamines (Berridge & Robinson, 1998). Perhaps elevated dopamine levels are one of the perks of dominance, rather than one of its precursors.
The relationships between behavioral styles and their physiological correlates evoke a question of the chicken and egg sort: What comes first, the physiological profile or the behavioral style that is associated with that profile? Do cortisol/serotonin levels themselves predispose an individual to a behavioral style, or is it possible that the behavioral style has other roots. If so, could it be that engagement in a particular behavioral style engenders the development of a particular physiological profile? As earlier noted, a variety of evidence indicates that these physiological profiles and their associated behavioral styles are reliably measured quite early in development, are stable over time, and are highly heritable (Sapolsky & Ray, 1989; Suomi, 1991). There is likely no clear answer to the chicken and egg question. The behavioral and physiological components of particular personalities are best thought of as co-regulated, that is, they arise from similar roots and once in place mutually maintain the links between them.
As for the social outcomes of this co-regulation, it should be emphasized again that neither physiological profile nor behavioral style invariably predicts social rank. Accordingly, the question of how personality maps on to social outcomes is not a simple one. There is evidence that experiential factors play a modulating role. Suomi (1997), for example, has shown in studies of cross-fostering that a rhesus macaque’s reactions in the absence of a stressor can be significantly affected by the parenting style of the adoptive mother. When stressed the adopted individual’s behavioral style is more predictable from the biological mother’s physiological profile and behavioral style. Additional work has shown that the social supports provided by members of a reactive monkey’s group can affect basal serotonin levels and the frequency of aggression, among other measures, thereby affecting the subsequent social rank of that animal (Barr et al., 2004). So, while physiological profile and behavioral style are linked and are often predictive of social outcome, experiential factors, particularly those related to parenting and social peer support, influence rank. There are different ways to become dominant, as there undoubtedly are different ways to become subordinate.
The presence of distinct personality profiles also evokes the adaptionist question: What are the advantages of evolving distinct behavioral styles and physiological profiles? In short, why are there different personalities? Adaptionist explanations, in the manner of Kipling’s "just so" stories, are all too easy to come by. The ease with which these explanations can be generated constitutes one of the weaknesses of evolutionary psychology. In the case of personality differences in primates one could easily fashion an explanation that involves minimizing direct competition among adults for resources by predisposing, though not inevitably determining individuals with certain physiological profiles and behavioral styles to certain social ranks. This type of mechanism may benefit individuals of any rank, at least in terms of the efficiency with which social rank and concomitant access to resources are established and maintained. This efficiency may have important advantages, especially within the type of complex societies that characterize many nonhuman primate species. It may also be that the long-term efficacy of a particular personality with respect to biological fitness may depend on the ecological context within which that personality is expressed. Impulsive animals might for example perform better than other animals when the risk of confronting predators is high, or when new habitats must be sought. If so, the presence of personality variation within a species makes considerable sense. (See Dingemanse & Réale, 2005, for a discussion of mechanisms that maintain variation within species.)
Interestingly, from this perspective personality differences may be more important in nonhuman primate societies than in human societies. In primate groups personality influences a number of aspects of daily life ranging from reactions to stress, interactions with peers and adults, access to resources, and even to an animal’s overall state of what may be referred to as subjective well being (Weiss et al., 2006). I imagine that the impact of personality differences in humans is not entirely dissimilar, but the benefits as well as the costs of particular personalities are surely mitigated by cultural practices. The idea is not that personality styles in humans have attained the same functional status as our appendix, but that personality differences are less attached to reproduction and survival in human cultures than in nonhuman societies. (It has been said of the human appendix that its major importance would appear to be financial support of the surgical profession, Romer & Parsons, 1986.) If so, an understanding of personality in nonhumans is important not simply because it sheds light on human nature, but because it sheds considerable light on nonhuman nature.
Do personality differences impact cognition?
For students of comparative cognition an understanding of personality in animals might be politely regarded as interesting in that it fleshes out our understanding of animal nature, and in doing so provides some interesting parallels with human nature. It may also be acknowledged that personality differences are one of the mechanisms than maintains behavioral variability, and, accordingly, they constitute one of the prerequisites for natural selection. It might be added though that an understanding of personality has little impact on the experimental study of cognition, which attempts to characterize the modal animal principally through the experimental study of group differences. As discussed earlier, Pavlov would have drawn the opposite conclusion, for he assumed that knowledge of these types helped predict the effectiveness of his experimental treatments.
Which position should be followed? The argument for the inclusion of personality differences in comparative cognition faces two obstacles. The first problem is whether Pavlov was indeed correct. Is there evidence that different personality styles predict differences in cognition? Even if the answer to this question is yes, there still remains the question of mechanics. How exactly would one take personality into consideration in studies of comparative cognition?
Pavlov aside, there is admittedly little evidence that individual differences in cognitive paradigms attach to differences in personality. Some dimensions of personality might seem more reasonably related to cognitive differences than are others. It is not surprising that differences in exploration are related to differences in learning. In a study involving female rhesus macaques, subjects were first screened for their exploratory ability by observing their interactions with a novel object placed in their home cage. Subjects were then exposed to an operant contingency that required touching another object in their home cage. Results revealed that a much higher percentage of exploratory subjects acquired the operant response compared to inhibited subjects (75% vs. 22%; Coleman et al., 2005).
There is other evidence that exploratory tendencies covary with learning differences (Matzel et. al., 2003). The connection between a trait like exploration and simple learning should not be surprising, particularly in procedures in which reactions to novelty and speed of conditioning were assessed using highly similar responses in the same context, as was the case in the study by Coleman et al. More compelling would be demonstrations that different personality styles covary with differences in cognition across a range of cognitive paradigms. That evidence is currently lacking, but it is not difficult to speculate—the adaptionist’s fallacy perhaps—that the personality styles identified in the study of primates might reasonably covary with differences in learning and memory. Impulsive monkeys, much like Pavlov’s Excitable type, may show deficits in procedures that require an interplay between excitation and inhibition, such as in differential reinforcement of low rate schedules, reverse contingency tasks (e.g., Boysen, Mukobi & Berntson, 1999), or tests of object permanence. Reactive monkeys might have difficulties in procedures where high anxiety impedes performance, or where exploration is required to contact the experimental contingencies. I imagine that this set of characteristics applies to many procedures ranging from aversive conditioning through spatial navigation. It is also the case that high arousal, either in the form of anxiety or excitability, has long been known to reduce cue utilization, the span of the attentional field (Easterbrook, 1959). In some cases this feature might be advantageous (e.g., reducing attention to irrelevant cues) but in other cases it may impair performance. This feature of arousal may be relevant to the performance of both impulsive and reactive animals in a variety of cognitive paradigms.
It should be said that some correspondences between cognition and personality have established neurobiological foundations. A number of studies have shown that the elevated cortisol levels characteristic of subordinate monkeys and a subset of dominant males negatively impact hippocampal functioning (e.g., Sapolsky, 2000). The hippocampus is rich in steroid receptors, and it normally serves a modulating function with respect to the release of glucocorticoids during stress. Ironically, prolonged stress has the effect of damaging the hippocampus and impairing precisely this function, thereby permitting elevated cortisol levels to persist, which in turn results in more hippocampal damage, and so on. It has long been known that hippocampal damage is associated with impairments in spatial navigation, memory, and attentional functioning (e.g., Landfield, Baskin, & Pitter, 1981; O’Keefe & Nadel, 1978). Additionally, in humans there is substantial evidence of correlations between high basal cortisol levels, hippocampal damage, and acceleration of the cognitive impairments characteristic of aging (Suomi, 1997). This same nexus between cortisol, hippocampal functioning, and cognition has also been observed in cases of severe depression and posttraumatic stress syndrome in humans, among other disorders.
It might be suggested that the importance of acknowledging the influences of personality on cognition depends on which sort of cognition is under consideration. If we are interested in what might be called physical cognition (e.g., tool use, spatial navigation, concept formation; Tomasello & Call, 1997), perhaps personality might be considered irrelevant. If our interest is more in social cognition (e.g., cooperative problem solving, alliance formation, imitation) then personality might assume greater importance given the reasonable assumption that different personality styles evolved principally to mediate the many demands attendant to social relations (Box, 1999). This dichotomy has common sense validity to it, but it also is flawed. The standard topics of physical cognition may be studied with nonsocial paradigms, but many if not all of them have commensurate social parallels. Transitive inference as an example has clear parallels to an individual animal’s understanding of social rank. Given that in some respects evolution is often conservative in the development of mechanisms—Why develop a new one when an old one might be modified?—it is likely that processes like transitive inference evolved to solve social as well as nonsocial problems. A similar argument suggests that the mechanisms mediating a primate’s ability to distinguish group members bear important similarities to the mechanisms found in humans by which the semantic meaning of a word or other symbol is inferred from experiencing that symbol in a variety of contexts. If so, these mechanisms may have contributed to the emergence of the semantic aspects of language (King, Rumbaugh, & Savage-Rumbaugh, 1999). In sum, if personality differences influence processes attendant to social relations, my guess is that they similarly affect nonsocial processes that present similar cognitive demands to the individual.
But does any feature of this argument matter in the pursuit of the goals of comparative cognition? If personality differences affect cognition at all they are expressed as part of the error term in standard group designs. They are part of the noise that constitutes the accepted price of using these designs. I do not want to belabor a point that is well understood by students of statistics, but it is of course true that the error term is a statement of our ignorance—we do not know why all that within group variance is occurring, but we trust that our experimental treatments will create substantially more variance between groups and thereby result in a significant effect.
We would be less ignorant if we could label a portion of that error variance and use it as a covariate. Even if one does not wish to study the relationships between personality and cognition directly, as did Pavlov, personality differences can still be used indirectly, that is, statistically. To the extent that personality differences affect cognition these differences can be used as a covariate, the variance owing to them extracted, thereby reducing the error term and affording the design greater precision. This point is well understood in the abstract, but unless I am mischaracterizing the available literature it is a technique seldom used in comparative cognition. An example of this possibility is presented in Appendix for the effects of context extinction on working memory. Without using the subject variable of activity (viz., low, medium, high) as a covariate the treatment effect is not statistically different than chance (p = .102). With the variance owing to activity statistically identified as a factor, the treatment effect is significant (p = .000).
A reasonable barrier to engaging in this type of strategy might be the time and effort involved in getting reliable personality ratings from subjects as a part of standard experiments. It may be easier than thought. With respect to using primates, personality assessments are typically made using established rating scales that can be administered in relatively short time periods (e.g., Bolig, Price, O’Neill, & Suomi, 1992; Stevenson-Hinde & Zunz, 1978). With rodents, quick assessments of activity level, exploration, and anxiety can be made using open fields, light-dark chambers, or elevated plus mazes. Protocols for these arenas are easily automated and can be completed in 5 minutes. Multiple animals can be run currently, thereby allowing a large number of animals to be screened efficiently prior to experimentation (e.g., Crawley, 2000; Locurto, Fortin, & Sullivan, 2003).
A related objection might be that in many studies of comparative cognition sample sizes are quite low owing to the rarity of a given species or its low representation in captivity. I imagine that this problem applies particularly to a number of primate species and to cetaceans. In these instances I would also make the case that it would be beneficial if some assessment of pre-experimental individual differences was made. It may be particularly useful to engage in this type of assessment in small-n designs given that in these designs extreme performance is not counterbalanced by a group average. If a given study uses, say, two rhesus macaques, and one of these animals happen to be reactive while the second one is more evenly-tempered, there is the chance that variations in their experimental outcomes will be due at least in part to pre-existing personality differences. The observed experimental differences between them may not be entirely cognitive per se, although they may be interpreted in that manner.
The fragile legacy of the two psychologies
The study of personality differences has its roots in a legacy different from the one that informs standard experimental designs. The study of individual differences, personality differences included, derives from correlational/descriptive approaches to data analysis. Experimental designs come from a legacy that attempts to determine causality through the manipulation of variables. Cronbach (1957) defined the correspondences between these two legacies in a classic article entitled "The two disciplines of scientific psychology." In it he argued that the two disciplines were essentially complementary: "The correlator’s mission is to observe and organize the data from Nature’s experiments. As a minimum outcome, such correlations improve immediate decisions and guide experimentation. At the best, a Newton, a Lyell, or a Darwin can align the correlations into a substantial theory." (p. 672).
Cronbach (1957, 1975) added that the separation between the two disciplines had impeded psychological research. Cross fertilization between the two disciplines would, he argued, permit an understanding of what he called aptitude x treatment interactions, cases in which subject characteristics influenced the outcome of experimental interventions. The point that Cronbach was pursuing here is relevant to the goals of comparative cognition. He was suggesting that these interactions are relevant for attempts at generalization, including attempts at modeling the performance of the modal subject.
I would add that both approaches are concerned with partitioning variance. Correlational approaches are interested in the variance shared or unique across different variables. Experimental approaches compare the variance created by manipulated treatments to the variance that is naturally occurring. Despite those differences the end point of both procedures is the identification of the sources of variance that attach to a given phenomenon. It is often said that correlational approaches are interested in description whereas experimental approaches are interested in explanation. I would argue that in a sense both procedures are ultimately concerned with prediction: Correlational techniques approach the problem of prediction via regression. Experimental techniques do so by direct manipulation of variables. If prediction is aided by an understanding of subject characteristics, and error variance is reduced as a consequence of that understanding, all the better for practitioners of either discipline.
Students of comparative cognition might be interested in Cronbach’s contention that historically the study of individual differences had been considered a fundamental aspect of comparative psychology. In the early 20th century comparative psychology included comparisons between individuals, species, cultures, and different development periods (see Locurto & Scanlon, 1998, for a similar discussion). Cronbach regretted the fact that while the other psychologies interested in individual differences (viz., personality, developmental) had remained "loosely federated" (p. 672), the animal behaviorists of this earlier era turned their discipline away from a truly comparative approach and into an experimental approach that focused on one or a few species. It is clear that the advantages of studying a variety of species have been appreciated in the modern study of comparative cognition. It is less clear that the advantages of including the correlational methodologies identified by Cronbach have been similarly appreciated.
References
Aiken, L. R. (1988). Psychological testing and assessment (6th ed.). Boston: Allyn & Bacon.
Barr, C. S., Newman, T. K., Shannon, C., Parker, C., Dvoskin, R. L., Becker, M. L., et al. (2004). Rearing conditions and rh5-HTTLPR interact to influence limbic-hypothalamic-pituitary-adrenal axis response to stress in infant macaques. Biological Psychiatry, 55, 733-738.
Berridge K., & Robinson, T. (1998). What is the role of dopamine in reward: hedonic impact, reward learning, or incentive salience? Brain Research and Brain Research Reviews, 28, 309-69.
Bolig, R., Price, C. S., O’Neill, P. L., & Suomi, S. J. (1992). Subjective assessment of reactivity level and personality traits of rhesus monkeys. International Journal of Primatology, 13, 287-306.
Box, H. (1999). Studies of temperament in simian primates with implications for socially-mediated learning. International Journal of Comparative Psychology, 12, 203-218.
Boysen, S. T., Mukobi, K. L., & Berntson, G. G. (1999). Overcoming response bias using symbolic representations of number by chimpanzees (Pan troglodytes). Animal Learning and Behavior, 27, 229-235.
Byrne, G., & Suomi, S. J. (2002). Cortisol reactivity and its relation to homecage behavior and personality ratings in tufted capuchin (Cebus apella) juveniles from birth to six years of age. Psychoneuroendocrinology, 27, 139-154.
Capitano, J. P., & Widaman, K. F. (2005). Confirmatory factor analysis of personality structure in adult male rhesus monkeys (Macaca mulatta). American Journal of Primatology, 65, 289-294.
Capitano, J. P. (1999). Personality dimensions in adult male rhesus macaques: Prediction of behaviors across time and situation. American Journal of Primatology, 47, 299-320.
Clarke, A. S., & Boinski, S. (1995). Temperament in nonhuman primates. American Journal of Primatology, 37, 103-125.
Coleman, K., Tully, L. A., & McMillan, J. L. (2005). Temperament correlates with training success in adult rhesus macaques. American Journal of Primatology, 65, 63-71.
Crawley, J. N. (2000). What’s wrong with my mouse? New York: Wiley-Liss.
Cronbach, L. J. (1957). The two disciplines of scientific psychology. American Psychologist, 12, 671-684.
Cronbach, L. J. (1975). Beyond the two disciplines of scientific psychology. American Psychologist, 30, 116-126.
Dingemanse, N. J., & Réale, D. (2005). Natural selection and animal personality. Behaviour, 142, 1159-1184.
Easterbrook, J. A. (1959). The effect of emotion on cue utilization and the organization of behavior. Psychological Review, 66, 183-201.
Eysenck, H. J. (1967). The biological basis of personality. Springfield, IL: Thomas.
Eysenck, H. J., & Eysenck, M. W. (1958). Personality and individual differences. New York: Plenum.
Fairbanks, L. A. (2001). Individual differences in response to a stranger: Social impulsivity as a dimension of temperament in vervet monkeys (ceropithecus aethiops sabaeus). Journal of Comparative Psychology, 115, 22-28.
Figueredo, A. J., Cox, R. L., & Rhine, R. J. (1995). A generalizability analysis of subjective personality assessments in the Stumptail Macaque and the Zebra Finch. Multivariate Behavioral Research, 30, 167-197.
Fox, N. A., Henderson, H. A., Marshall, K. E., & Ghera, M. M. (2005), Behavioral inhibition: Linking biology and behavior within a developmental framework. Annual Reviews of Psychology, 56, 235-262.
Goldberg, L. R. (1990). An alternative "description of personality": The big-five factor structure. Journal of Personality and Social Psychology, 59, 1216-1229.
Gosling, S. D. (2001). From mice to men: What can we learn about personality from animal research? Psychological Bulletin, 127, 45-86.
Gray, J. A. (1964). Pavlov’s typology. New York: Pergamon.
Higley, J. D., & Bennett, A. J. (1999). Central nervous system serotonin and personality as variables contributing to excessive alcohol consumption in non-human primates. Alcohol & Alcoholism, 34, 402-418.
Higley, J. D., & Suomi, S. J. (1989). Temperamental reactivity in non-human primates. In G. A. Kohnstamm, J. E. Bates, & M. K. Rothbart (Eds.), Temperament in childhood (pp. 153-167). New York: Wiley.
Itoh, K. (2002). Personality research with non-human primates: Theoretical formulation and methods. Primates, 43, 249-261.
Jones, A. C., & Gosling, S. D. (2005).Temperament and personality in dogs (Canis familiaris): A review and evaluation of past research. Applied Animal Behaviour Sciences, 95, 1-53.
Kagan, J., Saidman, N., & Arcus, D. M. (1992). Initial reactions to unfamiliarity. Current Directions in Psychological Science, 1, 171-174.
Kalin, N. H. (2003). Nonhuman primate studies of fear, anxiety, and temperament and the role of benzodiazepine receptors and GABA systems. Journal of Clinical Psychiatry, 64, 41-44.
Kaplan, J. R., Manuck, S. B., Fontenot, M. B., & Mann, J. J. (2002). Central nervous system monamine correlates of social dominance in Cynomolgus monkeys (Macaca fascicularis). Neuropsychopharmacology, 26, 430-443.
King, J. E., & Figueredo, A. J. (1997). The five-factor model plus dominance in chimpanzee personality. Journal of Research in Personality, 31, 257-271.
King, J. E., Rumbaugh, D. M., & Savage-Rumbaugh, E. S. (1999). Perception of personality traits and semantic learning in evolving hominids. In M. C. Corballis & S. E. G. Lea (Eds.), The descent of mind: Psychological perspectives on hominid evolution (pp. 98-115). New York: Oxford University Press.
Landfield, P. W., Baskin, R. W., & Pitter, R. A. (1981). Brain-aging correlates: Retardation by hormonal pharmacological treatments. Science, 214, 581-584.
Lilienfeld, S. O., Gershon, J., Duke, M., Marino, L., & de Waal, F. B. M. (1999). A preliminary investigation of the construct of psychopathic personality (psychopathy) in chimpanzees (Pan troglodytes). Journal of Comparative Psychology, 113, 365-375.
Locurto, C., & Scanlon, C. (1998). Individual differences and a spatial learning factor in two strains of mice (Mus musculus). Journal of Comparative Psychology, 112, 344-352.
Locurto, C., Fortin, E., & Sullivan, R. (2003). The structure of individual differences in heterogeneous strain mice across problem types and motivational conditions. Genes, Brain, & Behavior, 2, 1-16.
Mather, J. A., & Anderson, R. C. (1993). Personalities of Octopuses (Octopus rubescens). Journal of Comparative Psychology, 107, 336-340.
Matzel, L. D., Han, Y. R., Grossman, H., Karnik, M. S., Patel, D., Scott, N., et al. (2003). Individual differences in the expression of a "general" learning ability in mice. Journal of Neuroscience, 23, 6423-6433.
O’Keefe, J., & Nadel, L. (1978). The hippocampus as a cognitive map. Oxford: Clarendon Press.
Ozer, D. J. (1985). Correlation and the coefficient of determination. Psychological Bulletin, 97, 307-315.
Pavlov, I. P. (1906). The scientific investigation of the psychical faculties or processes in higher animals. Science, 24, 613-619.
Pavlov, I. P. (1941). Lectures on conditioned reflexes, vol. 2: Conditioned reflexes and psychiatry. W. Horsley Gantt, Translator. New York: International Publishers.
Raleigh, M. J., McGuire, M. T., Brammer, G. L., Pollack, D. B., & Yuwiler, A. (1991). Serotoninergic mechanisms promote dominance acquisition in adult male vervet monkeys. Brain Research, 559, 181-190.
Rang, H. P., Dale, M. M., Ritter, J. M., & Moore, P. K. (2003). Pharmacology (5th ed.). Edinburgh: Churchill Livingstone.
Ray, J. C., & Sapolsky, R. M. (1992). Styles of male social behavior and their endocrine correlates among high-ranking wild baboons. American Journal of Primatology, 28, 231-250.
Reif, A., & Lesch, K. P. (2003). Toward a molecular architecture of personality. Behavioural Brain Research, 139, 1-20.
Roeff, J. H., Sussman, R. W., & Strube, M. J. (2005). Personality traits in captive lion-tailed macaques (Macaca silenus). American Journal of Primatology, 67, 177-198.
Romer, A. S., & Parsons, T. S. (1986). The vertebrate body (6th ed.). Philedelphia, PA: Saunders College Pub.
Sapolsky, R. M. (2000). Glucocorticoids and hippocampal atrophy in neuropsychiatric disorders. Archives of General Psychiatry, 57, 925-935.
Sapolsky, R. M., & Ray, J. C. (1989). Styles of dominance and their endocrine correlates among wild olive baboons (Papio anubis). American Journal of Primatology, 18, 1-13.
Sapolsky, R. M. (1990). Andrenocortical function, social rank, and personality among wild baboons. Biological Psychiatry, 28, 862-878.
Seligman, M. E., & Weiss, J. M. (1980). Coping behavior: Learned helplessness, physiological change and learned inactivity. Behaviour Research and Therapy, 18, 459-512.
Sinn, D. L., & Moltschaniwskyj, N. A. (2005). Personality traits in Dumpling squid (Euprymna tasmanica): Context-specific traits and their correlation with biological characteristics. Journal of Comparative Psychology, 119, 99-110.
Stevenson-Hinde, J., & Zunz, M. (1978). Subjective assessment of individual rhesus monkeys. Primates, 19, 66-68.
Suomi, S. J. (1991). Uptight and laid-back monkeys: Individual differences in the response to social challenges. In S. E. Brauth, W. S. Hall, & R. J. Dooling (Eds.), Plasticity of development (pp. 27-56). Cambridge, MA: MIT Press.
Suomi, S. J. (1997). Early determinants of behaviour: Evidence from primate studies. British Medical Bulletin, 53, 170-184.
Tomasello, M., & Call, J. (1997). Primate cognition. New York: Oxford University Press.
Weiss, A., King, J. E., & Perkins, L. (2006). Personality and subjective well-being in Orangutans (Pongo pygmaeus and Pongo abelli). Journal of Personality and Social Psychology, 90, 501-511.
Yerkes, R. M. (1939). The life history and personality of the chimpanzee. American Naturalist, 73, 97-112.
Advantage of subject grouping
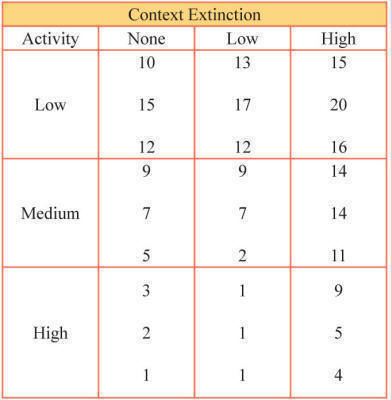
The example in this appendix illustrates the advantages and effects of using the subject variable of activity as a covariate. The experiment involves the effects of context extinction on working memory. The table below shows the scores of individual subjects as a function of context extinction (None, Low, High) and amount of activity (Low, Medium and High) with each cell having three scores. The data can now be analyzed in two ways.
Table 2A
 |
|
Data used in the analyses depicted in the following two figures. |
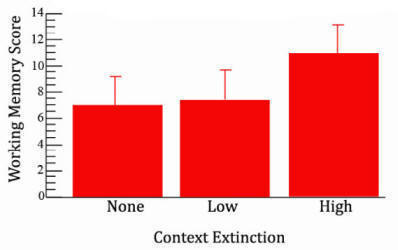
The next figure shows these data analyzed as a simple one-factor experiment without including subjects’ activity scores as a covariate. When grouped in this way the main effect of context extinction on the working memory score does not reach standard levels of significance. F(2,24)=2.51, p=.102, indicative of no effect of context extinction.
|
|
|
Figure 5A. Effect of context extinction without subject grouping. |
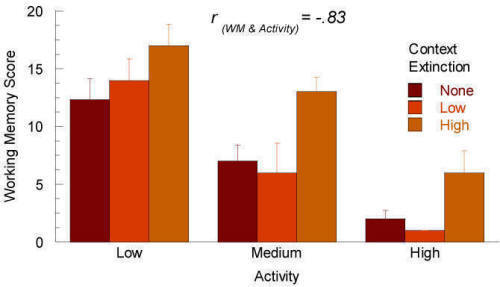
The next figure shows the same data grouped according to their activity scores. In this analysis, the variance owing to activity is now extracted from the data. When analyzed in this way the main effect of context extinction on the working memory score does reach significance, F(2,18)=13.66, p<.001, thus leading to a completely different experimental conclusion.
 |
|
Figure 6A. Subject variable of activity plotted against the main effect of context extinction. |
|
Preparation of this manuscript was supported by NSF grant BCS-0116089.
Correspondence concerning this article should be addressed to Charles Locurto, P. O. Box 75A, Department of Psychology, College of the Holy Cross, 1 College St., Worcester, MA 01610. E-mail: clocurto@holycross.edu |