| <<Vol. 1 Table of Contents | |
|
|
|
|
2006 |
Volume 1, pp 36-52 |

An Ontology for Comparative Cognition: A Functional Approach
Stan Franklin and Michael Ferkin
The University of Memphis
|
|
|||||||||||||||
|
|||||||||||||||
|
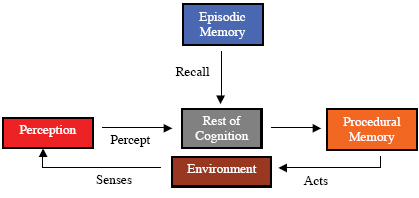
Introduction Comparative cognition, or the scientific study of how animals “think,” is rapidly becoming a scientific subfield of its own within Biology and Psychology. Scientists are becoming seriously concerned about cognitive processes in non-human animals. The study of cognition in animals has a long history beginning within the later nineteenth and early twentieth centuries with Darwin, Romanes, and Lloyd Morgan, and reaching our contemporaries (Balda, Pepperberg, & Kamil, 1998; Bekoff, Allen, & Burghardt, 2002; Blumberg & Wasserman, 1995; Griffin, 2001; Hauser, 2000; Shettleworth, 1998; Wasserman, 1993). A major thrust behind the interest in the study of intelligence in animals is the search for cognitive processes in the animal mind. Such an approach was initiated by the publication of the landmark volume, Cognitive Processes in Animal Behavior (Hulse, Fowler, & Honig,1978), and critically acclaimed books The Question of Animal Awareness, Animal Thinking, and Animal Minds (Griffin, 1976; 1984; 2001). Recent interest in understanding the minds of animals has shown a steady increase in articles appearing in journals such as Animal Cognition, Journal of Comparative Psychology and Journal of Experimental Psychology: Animal Behavior Processes, and an explosive growth in papers read at the annual meeting of the Comparative Cognition Society, now at 148 papers in 2005, up from 30 or so in 1993. A Google search on “comparative cognition” turned up over 10,000 hits. As with any developing scientific field, identifying and specifying the concepts that are important for the study, and their relations to one another, can be a major problem. In a recent review, Boysen and Himes (1999) stress that future research be conducted on conceptualization in non-human species, and that it should focus on imitation, tool use, mirror self-recognition and attribution of mental states. The use of terms for these concepts with different meanings from article to article causes difficulty in communication with readers and between scientists. For example, the term “cognitive” can be found used with a variety of different meanings in the various papers contained in a single anthology (Bekoff et al., 2002). Commonly accepted definitions, hopefully operational, are needed. Together, these accepted concepts and their relations would form a framework within which to study the subject, thereby creating an ontology of terms. We use the term ontology as a set of concept definitions with relations between them, specifying conceptualization within a system or discipline. This use of “ontology” is consistent with that used by computer scientists, but different from the meaning common among philosophers. In the case of comparative cognition, one might hope to import such an ontology from the study of human cognition, modifying it as needed. This approach, though a useful one, doesn’t solve the problem, because the issues of terms used with different meanings hasn’t yet been fully settled within the study of human cognition, itself a relatively young field (Eschenbach, Habel, Heydrich, & Rieser, 2003). Thus, in this paper we introduce a trial framework of concepts and their relations, an ontology, for the study of how animals may think. We hope that this paper will stimulate discussion leading to a more standard ontology of terms and important concepts for comparative cognition. We intend to err on the side of inclusivity, introducing more concepts and relations than are likely to survive a shakeout. We are seeking to provide a model and an ontology that revitalizes our approach to understanding how animal minds may work. One might view the model we describe below, and its ontology, as a kind of more detailed scientific paradigm in the sense of Kuhn (1970), or as a more detailed central core of a research program in the sense of Lakotos (1974). Taking the latter view, we hope that our model and ontology will provide a central core by providing “definitions and assumptions that coherently map out directions for research” (Kamil, 1998). The framework of the model and the ontology that we introduce is based on the IDA (Intelligent Distribution Agent) model of cognition (Baars & Franklin, 2003; Franklin, 2001b; Franklin, 2005a; Franklin, Baars, Ramamurthy, & Ventura, 2005), a computational and conceptual model derived from a working software agent (Franklin, 2001a) that assigns new jobs to sailors at the end of their tour of duty (see below). The IDA model is a kind of “cognitive theory of everything,” including perception, feelings and emotions, various kinds of memory, consciousness (attention), several kinds of learning, action selection, automization, deliberation, volition, metacognition, etc. In the section that follows we introduce the proposed framework piece by piece, based on functional needs of animals (and other agents). Wherever possible, we will refer the reader to the existing scientific literature on human and animal cognition concerning the concept at hand. For the convenience of the reader, each newly introduced term in the proposed ontology will be highlighted and included in an appended glossary. The glossary will also contain needed technical terms (un-highlighted) that are not part of the ontology. Later sections will describe the IDA model and its cognitive cycle, while carrying along an example scenario. We’ll close with a brief discussion of the strengths and weaknesses of the IDA model and our derived ontology. Agents, Animals & Cognitive Functionality Agents, Action Selection and Cognition An autonomous agent (Franklin & Graesser, 1997) is a system situated in, and part of, an environment, which senses that environment, and acts on it, over time, in pursuit of its own agenda. Biological examples of autonomous agents include humans and other animals. Non-biological examples include some mobile robots, and various computational agents, including artificial life agents (Langston, 1989), software agents (Franklin & Graesser, 1997) and many computer viruses. We’ll often use “agent” as an abbreviation for “autonomous agent.” In biological agents, the agenda arises from evolved drives and their associated goals; in artificial agents, the agenda arises from drives and goals built in by their designers. Such drives, like the drive to mate in rodents, act as motive generators (Sloman, 1987), and must be present, whether explicitly represented, expressed causally, or implemented by feelings and emotions as in humans and other animals (Franklin & McCauley, 2004; Panksepp, 1998). Every autonomous agent also acts in such a way as to possibly influence what it senses at a later time. In other words, it is structurally coupled to its environment (Maturana, 1975; Maturana, & Varela, 1980). We’ll be concerned with animals, including humans, thought of as autonomous agents, situated in their environments, sensing their environments and acting on their environments (Figure 1).
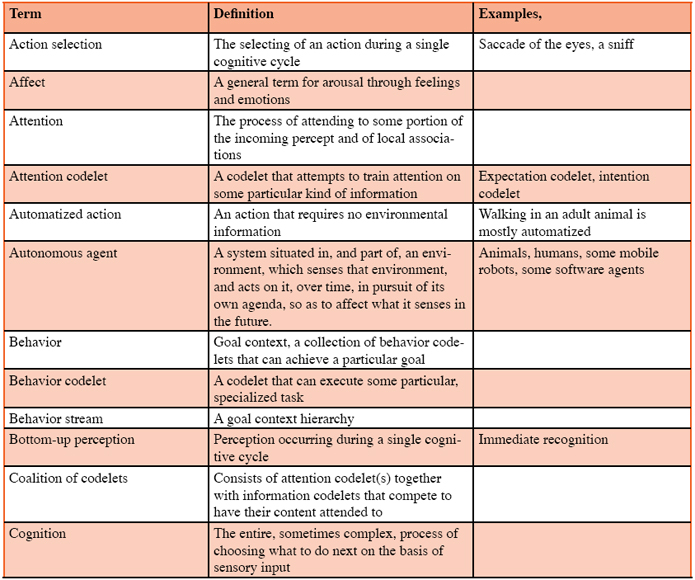
Every autonomous agent, including humans and other animals, spends it waking life in the moment-to-moment responding to the only question there is: “What shall I do next?” (See the Action Selection Paradigm in Franklin, 1995) Thus, this deciding what to do next constitutes the major activity of any agent between each sensing of its environment (sniff, glance, etc.) and the agent’s next action upon it. Freeman (1999) refers to this process as the forming of intentions. Note that most of these actions in animals consist of directing sense organs via a sniff, a saccade of the eyes, a perk or directing of an ear, a turning of the head, a sound production for echolocation, etc. Using the term in an unusually broad sense, we shall refer to this entire, cyclic, often complex, process of choosing what to do next based on sensing the current environment and upon current goals as cognition (Figure 2). This is a broader than usual use of the term since psychologists often exclude perceptions and emotions from cognition (Dalgleish & Power, 1999). For example, Pylyshyn (1999) describes a controversy over the relationship of visual perception and cognition, using the narrower concept of cognition. In the following sections we will partition cognition into interacting functional modules, gradually building an ontology for the scientific study of comparative cognition. Perception Current scientific literature mostly discusses perception from the point of view of psychophysics (Hirsh,1996) or neurobiology (Lu & Sperlin, 2001). Here we take a different point of view. Not only must every animal (autonomous agent) sense its environment, it must “make sense” of what it senses. We will refer to this process of assigning meaning to incoming sensory data as perception. Perception certainly depends upon the sensitivity of the agent’s sensory apparatus to sensory input, but is also a productive function. While light of a certain wavelength reflects off objects in the world, the color red exists, if it does, only by means of the perception of an animal (Oyama, 1985). Perception produces red. A tree falling in the forest will produce waves in the air, but no sound unless an animal hears it. There are molecules of certain substances in the air, but they are not perceived as chemical signals, unless an animal encounters them (Ferkin & Johnston, 1995a; b; Ferkin, Lee, & Leonard, 2004a; Ferkin, Li, & Leonard, 2004b). This is how Christof Koch (2004a) describes the perception of color: The much-cherished sense of color is a construct of the nervous system computed by comparing activity in the different cone classes. There are no “red” or “blue” objects in the world. Light sources, such as the sun, emit electromagnetic waves over a broad wavelength spectrum. Surfaces reflect this radiation over a continuous range and the brightness incident to the eyes is continuous as well. Nevertheless, all of us persist in labeling objects as read, blue, violet, purple, magenta, and so on. Color is not a direct physical quantity, as is depth or wavelength, but a synthetic one. Different species have fewer or more cone types, and therefore experience quite different colors for the same objects. For example, some shrimp have eleven cone classes. Their world must be a riot of colors! (Koch, 2004, page 52.) Perception can be bottom-up, that is, occurring during a single sense –> cognize –> act cycle, or top-down, that is requiring multiple cycles. Top-down perception may utilize new sensory data provided as a result of an action, for example the turning of the head. It may be helped by input from various forms of memory (see below).
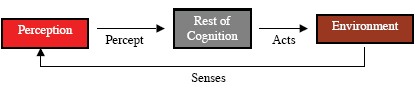
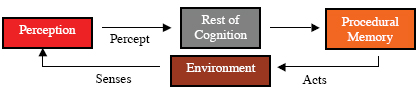
Perception assigns meaning to sense data, that is, it interprets the data so as to make it useful to the animal (Glenberg, 1997). This interpretation may include the identification of individual objects, including other animals, or categorization of objects, or relations between them, etc. The meaning assigned to the sense data of a particular cognitive cycle is the percept of that cycle (Figure 3). A collection of sensory features arising from a large fanged animal may be identified as an object. This object may be categorized as a predator. Both the identification and the categorization might be parts of the meaning assigned, that is, part of the percept. Since much of the sense data of animals, and some artificial agents, is irrelevant to the needs of the animal, perception often acts also as a filter, with only relevant portions of the meaning becoming part of the percept. The meaning of “meaning” is itself a complex issue involving such philosophical concerns as intentionality (e.g. Searle, 1983) and symbol grounding (e.g. Harnad,1990), as well as neurobiological issues such as binding (e.g. Roskies,1999) and computational issues such as representation (e.g. Davis, Shrober, & Szolovits, 1993). A full discussion of these issues is beyond the scope of this paper. Suffice it to say here that we take the viewpoint of situated or embodied cognition with respect to these issues (Anderson, 2003). Memory In the existing psychological and neuroscience literatures, one of the most confusing concepts is that of memory. Memory includes dozens of variants, many of which are not carefully distinguished from other variants. These variants are demarked by such adjectives as associative, autobiographical, conceptual, declarative, episodic, exemplar, implicit, long-term, long-term working, procedural, prospective, recall, recognition, semantic, sensory, short-term, spatial, working, and no doubt many others. Memory systems are most often categorized by their duration or their function. Attempts at explication can be found elsewhere (Franklin et al., 2005; Schacter & Tulving, 1994). Here we must limit ourselves to a discussion of the few of these variants that the authors consider to be central to our ontology and to our model. Cognition for animals seems often to include some form(s) of memory. Perception is a form of memory in that not only are existing meanings assigned, objects recognized and categorized, etc., but also new meanings are created. New objects, categories, and relationships are learned. In the psychological literature, perception is most often not considered as a form of memory (but see Rauchs, Desgranges, Foret, & Eustache, 2005). Perceptual learning seems to be ubiquitous among animals (Papaj & Lewis, 1993). For example, meadow voles can distinguish between unfamiliar and familiar conspecifics, littermates and non-littermates, and between sexually receptive and sexually quiescent opposite-sex conspecifics. Meadow voles respond preferentially to the odors of littermates relative to non-littermates by spending more time investigating the odors of the former as compared to those of the latter (Ferkin 1990; Ferkin, Tamarin, & Pugh, 1992). Adult female voles behave amicably towards familiar females but not towards unfamiliar females, whereas adult male voles behave agonistically towards familiar males but not unfamiliar males (Ferkin, 1988). Male voles over-mark the scent marks of females in heightened sexual receptivity, during postpartum estrous, as compared to those of females that in other states of sexual receptivity (Ferkin et al., 2004a; b) Depending on the social context, the perceptual memory of voles may last several hours to several days (Ferkin, 1992). Perceptual memory can be fleeting or long-term. For instance, a new person met briefly at a party may not be recognized a few weeks later, while a friend from childhood who hasn’t been seen for decades may be recognized in spite of the changes brought by age. The intervening steps between perception and perceptual learning, that is, the learning mechanism, will be clarified during the discussion of the IDA cognitive cycle below. Many animals also learn to perform new tasks, motor behaviors, or to improve their performance of existing behaviors. Procedural memory stores the procedures for executing these behaviors (Figure 4). A newborn ungulate, say a wildebeest, on a savannah takes a few minutes to learn to stand, walk, and run. Edelman (1987) describes an initial, primary repertoire of such procedures (neuronal groups) with which an animal is born, as well as a secondary, learned, repertoire including more complex behaviors (See Figure 4).
Many, but we suspect not all, animals have some form of episodic-like memory, that is, an associative, content-addressable, memory for events, for the what, the where and the when (Clayton, Busey, & Dickinson, 2003; Griffiths, Dickinson, & Clayton, 1999). Memories of events are recovered by means of their associations with cues, the content. (See Step 4 of the IDA cognitive cycle below.) Wishing to avoid entanglement fruitless debates about mental images in non-human animals, animal behavior researchers have spoken of episodic-like memory. Episodic-like memory as a memory about things that happened at a particular time and place, or the “what, where, and when” (Clayton & Dickinson, 1998; Clayton, Griffiths, & Dickinson, 2001; Griffiths et al., 1999). These memories must be based on a one-time experience, to preclude learning or conditioning, and must be stored in long-term memory. The most convincing studies testing episodic-like memory in animals comes from work on food caching in scrub jays. The major finding of this work is that scrub jays remember not only what food item they cashed and where, but when. Moreover, the scrub jays adjusted their caching behavior in anticipation of future events (Clayton & Dickinson,1998; Clayton et al., 2001; Emery & Clayton, 2001). The researchers concluded that episodic-like memories consist of spatio-temporal location of food items based on a single caching experience and that animals may recall the past and plan for the future (Clayton et al., 2003). Functionally, episodic and episodic-like memories are the same, though the underlying mechanism of the former would require consciousness (Tulving, 2001). In humans, episodic memories are typically recalled by means of mental images, including visual images, auditory images, olfactory images, etc. Neural apparatus in the human perceptual systems are reused for this purpose (Baddeley, Conway, & Aggelton, 2001; O’Craven & Kanwisher, 2000).
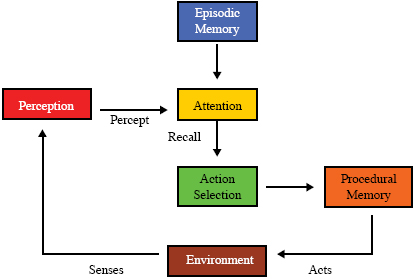
An example will help to distinguish episodic from perceptual memory. The feeding behavior of rats has been studied in an eight-arm radial arm maze (Olton & Samuelson, 1976). With four arms baited and four not (with none restocked), normal rats learn to recognize which arms to search (perceptual memory). They also remember which arms they have already fed in on this particular day (episodic memory) so as not to search there a second time. Rats with their hippocampal systems excised still learn to recognize the baited arms, but cannot remember which of the baited arms they have already fed in. They lose their episodic memory but retain perceptual memory. Responding to cues from perception, episodic memory provides information from prior events to the rest of cognition. A reuse of perception may be required to interpret the information recalled from episodic memory (Figure 5). Episodic memories may be transient, that is, with fairly rapid decay, or long-term, that is, capable of holding some memories perhaps indefinitely. Perceptual, episodic and procedural memory, depend to some extent, and in different ways, on association. In perceptual memory an object is associated with its features, a category with its members. Recall from episodic memory is accomplished in animals (and in at least some artificial agents) by means of associations with a cue. Improvement of performance during procedural learning is accomplished in animals by associating particular actions with desired results. Thus association plays different roles in the various memory system and their various forms of learning, and can be expected to require distinct mechanisms. Attention and Action Selection
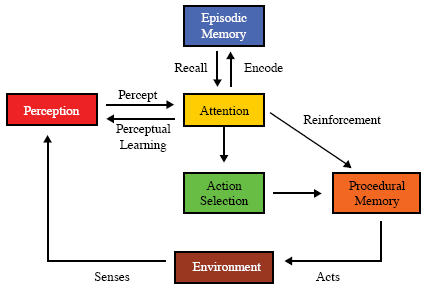
In most, if not all, animals, the richness of sensory perception, together with retrieved information from any episodic memory it might have, vastly complicates the process of deciding what to do next, that is, its action selection process. Some means is needed for deciding what parts of this vast array of information is currently relevant and should be attended to when selecting the next action. A competitive attention mechanism solves this relevance problem, providing relevant, current information to the action selection process (Figure 6). In bottom-up perception during a single cycle (see page 5) perceptual activity is pre-attentive. Perception acts as one filter, attention as a subsequent filter, both in the same cycle. In higher-level top-down perception requiring multiple cycles, attention during previous cycles affects current perception. There’s a vast psychological and neuroscience literature on attention and action selection (e.g., Perr & Hodges, 1999; Proctor, Wang, & Vu, 2002). Below we’ll see how the global workspace theory of consciousness and cognition accounts for animals solving this relevance problem (Baars, 1988; 2002). Learning As mentioned earlier, perceptual learning seems ubiquitous among animals. Procedural learning, learning to perform new tasks e.g. the standing, walking, running mentioned above, or to improve the performance of existing behaviors also seems widespread, at least among mammals and birds. Often called motor learning, procedural learning has been defined as “ a set of [internal] processes associated with practice or experience leading to relatively permanent changes in the capability for responding” (Schmidt, 1988 p. 346). There’s a huge psychological literature on procedural learning, much of it concerning testing human subjects learning such sensory-motor tasks as the pursuit rotor task under various laboratory conditions (e.g. Wulf, Hoss, & Prinz, 2001). A typical such task is the pursuit rotor task in which the subject learns to track a metal dot on a turntable platter with a flexible metal rod. Such tasks have been studied in the context of the effect of practice (Brown & Bennett, 2002), of sleep deprivation (Smith & MacNeill, 1994) and of various mental illnesses (e.g. van Gorp, Altshuler, Theberge, & Mintz, 1999). Some animals also display episodic-like learning, the encoding of events in episodic memory, or episodic- like memory, for example, caching birds remembering locations (Clayton & Dickinson, 1998; Clayton et al., 2001; Griffiths et al., 1999). Amnesiac patients, such as the famous HM, demonstrate the distinct neural mechanisms required for episodic learning, of which they are incapable, and procedural learning, where they perform normally (Corkin, 1968). Olton and Samuelson’s (1976) experiment with rats searching for food in an eight armed maze demonstrates the same distinction.
Animals learn only what they attend to. Thus, short-term primings, resulting from stimuli of which the human subject is not conscious, constitute a trivial exception to this assertion (Eimer & Schlagecken, 2003). Attention produces perceptual learning, the encoding of events in episodic memory (in some animals), and the reinforcement for procedural learning (Figure 7). Note that reinforcement, a mechanism for procedural learning, is always ultimately based internally on pleasure or pain, that is, on positive or negative affect. We now have the beginnings of a functional ontology for the scientific study of comparative cognition; the initial steps in that only functionality within a single cognitive cycle is considered. But, many cognitive processes require multiple cycles. On example of such a multi-cyclic process is social learning via imitation, which occurs in humans and some other animals. In the next section we’ll take up a few of these multi-cyclic processes. Intentions, Goals, Volition and Actions Actions require motivation. Tautologically, without motivation no action would be taken. We’ve previously talked about drives as primary motivators. Goals, intentions to restructure the environment, or an agent’s relation to it, in particular ways, are in the service of other goals and, ultimately, of one or more drives. Volition refers to the process of arriving at a goal. An intention arises in an animal (agent) when it decides to achieve a particular goal. Neuroscientist Walter Freeman puts intentionality as the corner stone of his system, and claims that almost all of the brain of an animal is directly involved in arriving at any intention (Freeman, 1999). Please be aware that philosophers use the term “intentionality” with and entirely different meaning. To them it designates the ability to be about something, to represent something, to stand for something (Dennett, 1987). Note that human level consciousness is not required for intention. Actions are undertaken in pursuit of goals and in response to intentions. Such actions may be informationally mediated or they may be automatized. An informationally mediated action requires information from the environment for its enactment. A predator’s pursuit of a prey is informationally mediated since it must continually update its information as to the prey’s rapidly changing location. In contrast, the predator’s actions in running are, for the most part, automatized in that they require only occasional information from the environment (Mounoud, 1993). An animal’s ongoing activity can be seen as very occasional acts of volition (check readiness to mate) among numerous informationally mediated actions (avoid that bush, move behind her, sniff), interspersed with automatized actions (walk) (turn head). Once the goal of checking readiness to mate is chosen, ‘avoid that bush’ is selected in the service of that goal, requiring information from the environment, but no further act of volition. The walking is mostly automatized with occasional sensing of the environment to avoid unwanted collisions. This volition/action process can occur in humans, and possibly in other animals, at three different levels of intricacy as described by Sloman (1999). In a deliberative process, options may be constructed and evaluated, and plans created. This process can be thought of as using imagination, an internal simulation of interaction with the environment (O’Connor &Aardema, 2005; Shanahan, 2005), in the service of decision making, problem solving or planning. Once again, there is a large psychological literature devoted to this topic under various names (Busemeyer, Medin, & Hastie, 1995; Kahneman, Slovic, & Tversky, 1982). A likely animal example of deliberation is the Portia fimbriata jumping spider stalking its prey by making a lengthy detour around and above the prey, and loosing sight of the prey for a significant time period, before sighting the prey again and descending on it from above (Wilcox & Jackson, 1998). Almost all volition and action selection in animals, including humans, result from reactive processes, that is processes that are not deliberative. Note that reactive processes may vary from quite simple, such as reflexes, to extraordinarily complex involving sophisticated perception, memory and action selection, such as those described in the IDA cognitive cycle (see below). In addition, we suggest that such deliberative processes depend on underlying reactive processes. A third level of such processing, metacognitive processing, is described in humans as thinking about thinking (Hacker & Bol, 2004; O’Connor & Aardema, 2005). Sloman (1999) refers to this term as meta-management or reflexive. Metacognition is the term favored by psychologists. For example, a father may metacognitively decide that in a recent interaction with his daughter, he was too hard on her, and resolves to be more understanding. Do metacognitive processes occur in non-human animals? Experimental data suggests that some non-human animals have “functional features of or parallels to human metacognition and human conscious cognition” (Smith & Washburn, 2005). Intermediate Summary In summary, we’ve described in the previous sections the concepts (terms) in the proposed functional ontology for comparative cognition, together with a skeletal outline of their relations and interactions, and example references to the empirical literatures of these subjects. This provisional ontology was derived primarily from functional consideration based upon the definition of autonomous agent. Much is left to do. We must: 1. Flesh out these relations and interactions between the concepts of the ontology.The IDA model and its cognitive cycle, which motivated our ontology, will help with at least tasks 1, 2, and 4 above. A superficial account of some of the computational mechanisms may also prove useful. In the next section we discuss the IDA model, and its cognitive cycle, together with a minimal computational account. The IDA Model and its Cognitive Cycle As discussed above, our functional ontology for comparative cognition is based on and motivated by the IDA model of human cognition, which we will describe in this section along with its cognitive cycle. IDA provides a conceptual (and computational) model of cognition (Franklin, 2001b; 2005a) partially implemented as a software agent (Franklin & Graesser, 1997). The IDA model implements and fleshes out Global Workspace theory (GWT) (Baars, 1988; 2002), which suggests that conscious events involve widespread distribution of focal information needed to recruit neuronal resources for problem solving. As we’ll see in the middle steps of the cognitive cycle below, the role of consciousness according to GWT is to select the most relevant information in the current situation, and to broadcast it widely within the cognitive system (brain) in order to recruit internal resources that can make a useful response. Those middle steps in the cognitive cycle below describe how this is done. The IDA implementation of GWT yields a fine-grained functional account of the steps involved in perception, several kinds of memory, “consciousness,” context setting, and action selection. We make no claim of phenomenal (subjective) consciousness for software agents, but without opposing empirical evidence, we are not willing to reject the view that it may be involved in some cognitive processes for some non-human animals under particular conditions (Franklin, 2005b; Griffin, 1976; 1984; 2001; Hauser, 2000; Merker, 2005). We use the term “consciousness” to refer to functional consciousness, essentially the recognition of the thinking subject of its own actions, which may or may not be dependent on the subject’s thoughts or feelings that motivate them. Cognitive processing in IDA consists of continually repeated traversals through the steps of a cognitive cycle (Baars & Franklin, 2003; Franklin et al., 2005), as justified above (see text surrounding Figure 1.) and described below. The IDA architecture includes modules for perception (Zhang, Franklin, Olde, Wan, & Graesser, 1998), various types of memory (Anwar & Franklin, 2003, Franklin et al., 2005), “consciousness” (Bogner, Ramamurthy, & Franklin 2000), action selection (Negatu & Franklin, 2002), deliberation, and volition (Franklin, 2000). The mechanisms of these modules are derived from several different “new AI” sources (Hofstadter & Mitchell, 1995; Jackson, 1987; Kanerva, 1988; Maes, 1989). In addition to the computational model, we will also speak of the conceptual IDA model, which includes additional capabilities that have been designed but not implemented, including mechanisms for feelings and emotions, and mechanisms for perceptual and procedural learning. The IDA conceptual model contains several different memory systems. Perceptual memory enables identification, recognition and categorization, including of feelings, as well as relationships and situations. Working memory provides preconscious buffers as a workspace for internal activities. Please note that this usage of the term “working memory” differs from that common in the psychological literature, where conscious access is assumed (Baddeley, 1993), We must deal with the preconscious buffers of working memory due to the finer temporal grain size of the IDA cognitive cycle (see below). Procedural memory is long-term memory for skills. As discussed above, episodic memory is memory for events, the “what the where and the when.” In the IDA model, transient episodic memory is a content-addressable associative memory with a moderately fast decay rate (Conway, 2002; Franklin et al., 2005). It is to be distinguished from long-term episodic memory, which is often called declarative memory as we will below (Franklin et al., 2005). Some events may decay rapidly from declarative memory, while others may last a lifetime. Declarative memory is often subdivided into autobiographical memory for complete events, and semantic memory containing facts, for example “Paris is the capital of France.” Semantic memory is thought to contain events that retain their what, but have lost there where and when. It’s not clear to what extent, if any, non-human animals have semantic memory. Next we describe the conceptual IDA’s cognitive cycle, most, but not all, of which, has been implemented. In several of the steps of the cognitive cycle we refer to codelets, which correspond to the processors of GWT. A codelet is a special purpose, relatively independent, mini-agent typically implemented as a small piece of code running as a separate thread in the computational IDA. (Since a codelet is an implementational concept in IDA, we do not include it in our ontology for comparative cognition.) We hypothesize that cognitive cycles occur five to ten times a second in humans, cascading so that some of the steps in adjacent cycles occur in parallel (Baars & Franklin, 2003; Franklin et al., 2005). Seriality is preserved in the conscious broadcasts. We propose that every animal lives its waking life in a continual cascade of such cognitive cycles similar to the one we describe next, but perhaps missing some of the steps. In what follows we will describe each of the steps in IDA’s cognitive cycle, stated as if applying to a human, while also carrying along their application in the mind of a hypothetical male vole in highlighted text. Suppose our male vole has just turned a corner, and encountered scent marks from many different conspecifics (Ferkin & Johnston, 1995a). Some of these scent marks are old and some are fresh, some are overlapping and some are not. Our male is capable of detecting these many marks, identifying the donor, and identifying the identity of the donors that deposited the most and the freshest marks (Ferkin, Dunsavage, & Johnston, 1999; Ferkin, Mech, & Paz-y-Mino, 2001; Ferkin et al., 2004a; b; Ferkin, Pierce, Sealand, & delBarco-Trillo, 2005). He then is able to distinguish between the different scent donors and respond accordingly to the donor are of most interest to him; this donor would likely be a sexually receptive female with whom he would attempt to copulate (delBarco-Trillo & Ferkin, 2004). Our vole’s discrimination between the different scent donors would have involved perceptual learning. Keep in mind that the cognitive cycle to be described takes, in total, only a fifth of a second or so to complete. Also note that our description of the IDA cognitive cycle will refer to mechanism details such as codelets, the slipnet and others that are part of our IDA implementation, but are not germane to the functional ontology we detailed above. We include this description of the IDA cognitive cycle to show the reader the origin of the suggested ontology, and also as an example of how a computational model can be built using this ontology as a framework. In the appended glossary, such implementation mechanisms are included for the convenience of the reader, but are not highlighted. Highlighting is reserved for concepts that are part of the functional ontology. For more detail on these implementation mechanisms please consult the cited references. The first step of the IDA cognitive cycle fleshes out the
material in the Perception section above. a. Early perception: Input arrives through senses. Specialized perception codelets descend on the input. Those that find features relevant to their specialty activate appropriate nodes in the slipnet (a semantic net with activation) (Franklin, 2005c). This perceptual memory system identifies pertinent feeling/ emotions along with objects, categories and their relations. Feeling nodes for interest and for sexual arousal are somewhat activated. Suppose all of these activated nodes are over threshold and become part of the percept. Steps 2 and 3 of the IDA cognitive cycle implement most of the concepts discussed in the section on Memory above. 2. Percept to Preconscious Buffer. The percept, including some of the data plus the meaning, is stored in preconscious buffers of IDA’s working memory. In humans, these buffers may involve visuo-spatial, phonological, and other kinds of information. Feelings/emotions are part of the preconscious percept. The percept here has identified the freshest scent marks coming from a female in postpartum estrus. These females readily mate when they encounter males, however, they are only receptive to males for 12 hours after they deliver pups (Ferkin et al., 2004a). 3. Local Associations. Using the incoming percept and the residual contents of the preconscious buffers (content from precious cycles not yet decayed away), including emotional content, as cues, local associations are automatically retrieved from transient episodic memory (TEM) and from declarative memory. The contents of the preconscious buffers, together with the retrieved local associations from TEM and declarative memory, roughly correspond to Ericsson and Kintsch’s (1995) long-term working memory and to Baddeley’s (2000) episodic buffer. These local associations include records of the agent’s past feelings/emotions, and actions, in associated situations. Assuming that our male vole possesses declarative memory, the retrieved local associations may include the memory of a previous sexual encounter with this particular female and his reaction to it. For example, our male vole may have a memory of this female, when she was not in postpartum estrus, but simply pregnant and not sexually receptive (Ferkin & Johnston, 1995a; b), which allows our male vole to anticipate that this female will only be in postpartum estrus for a few hours, and then she becomes not interested in mating. Although such expectation may come from either perceptual memory or semantic memory, anticipating the what (a female is highly sexually receptive for a relatively narrow window), the when (a female may no longer be highly sexually receptive), and the where (the location of that female relative to other female voles in the area), suggest that such processing may involve an episodic–like memory. The remaining steps of the IDA cognitive cycle flesh out the contents of the sections above on Attention and Action Selection, and on Learning. 4. Competition for Attention. Attention codelets view long-term working memory, and bring relevant, important, urgent, or insistent events to consciousness. (Consciousness here is required only in the functional sense as defined in Global Workspace Theory (see above) and as defined by its role in the middle steps of this cognitive cycle.) Some of them gather information codelets (codelets that carry content), form coalitions and actively compete for access to consciousness. The coalition with the highest average activation over its codelets wins the competition. The competition may also include such coalitions from a recently previous cognitive cycle. Present and past feelings/ emotions influence this competition for consciousness. Strong affective content strengthens a coalition’s chances of being attended to (Franklin & McCauley, 2004). An attention codelet that is on the lookout for sexual opportunities will form a coalition with information codelets carrying the other vole’s identity, her reproductive status and readiness to mate, some details of the previous encounter, and the feelings associated with the current percept and the previous encounter. This coalition will compete with other such for “consciousness,” but may not win the competition. Suppose our male’s first encounter with that female’s odor indicated that she has also attracted the attention of a predator, (fresh weasel scent marks are present), which has also become part of the percept, along with a strong fear. In this case, the coalition formed by an attention codelet on the lookout for danger may well win the competition, and the male voles may not respond by seeking out this female. 5. Broadcast of Conscious Contents. A coalition of codelets, typically an attention codelet and its covey of related information codelets carrying content, gains access to the global workspace and has its contents broadcast to all the other codelets in the system. In humans, this broadcast is hypothesized to correspond to phenomenal consciousness. The conscious broadcast contains the entire content of consciousness including the affective portions. Suppose there were no predator and that the coalition about the female vole was attended to, that is, it came to the male vole’s “consciousness.” Several types of learning occur. The contents of perceptual memory are updated in light of the current contents of consciousness, including feelings/emotions, as well as objects, categories and relations. The stronger the affect, the stronger the encoding is in memory. Possibly along with others, the nodes in perceptual memory for the particular female vole, for the category of female voles, for readiness to mate, and for sexual interest would each have their base-level activations strengthened. Transient episodic memory is also updated with the current contents of consciousness, including feelings/emotions, as events. The stronger the affect, the stronger would be the encoding in memory. (At recurring times not part of a cognitive cycle, the contents of transient episodic memory are consolidated into long-term declarative memory.) If our male vole possesses a transient episodic memory, the event of having again encountered this particular female vole, her condition, and his reaction to her would be encoded, taking information from the “conscious” broadcast. Procedural memory (recent actions) is updated (reinforced) with the strength of the reinforcement influenced by the strength of the affect. The prior acts of turning the corner and sniffing the encountered scent marks would be reinforced. In this case, both acts would have been so over learned that their base-level activations would have been saturated, so that the reinforcement would have little or no effect. Thus, perceptual, episodic and procedural learning occur with the broadcast in each cycle. 6. Recruitment of Resources. Relevant behavior codelets respond to the conscious broadcast. These are typically codelets whose variables can be bound from information in the conscious broadcast. If the successful attention codelet was an expectation codelet (see Step 9 below) calling attention to an unexpected result from a previous action, the responding codelets may be those that can help to rectify the unexpected situation. Thus consciousness solves the relevancy problem in recruiting internal resources with which to deal with the current situation. The affective content (feelings/emotions), together with the cognitive content, helps to attract relevant resources (behavior codelets, processors, neural assemblies) with which to deal with the current situation. Possibly among others, behavior codelets for turning the head, for turning the body, for sniffing the scent marks and for moving in the direction that the female vole was traveling, binding such variables as the direction of the head and the direction of body and motion from the information in the broadcast. 7. Setting Goal Context Hierarchy. The recruited processors (behavior codelets) use the contents of consciousness, including feelings/emotions, to instantiate new goal context hierarchies, bind their variables, and increase their activation. Goal contexts are potential goals, each consisting of a coalition of procedures (behavior codelets), which, together, could accomplish the goal. Goal context hierarchies can be thought of as high-level, partial plans of actions. It is here that feelings and emotions most directly implement motivations by helping to instantiate and activate goal contexts, and by determining which terminal goal contexts receive activation. Other, environmental, conditions determine which of the earlier goal contexts receive additional activation. In this case a goal context hierarchy (behavior stream) to seek out the female vole would likely be instantiated, activated, and some of its variables bound with information from the broadcast. 8. Action Chosen. The behavior net chooses a single behavior (goal context), perhaps from a just instantiated behavior stream or possibly from a previously active stream. This selection is heavily influenced by activation passed to various behaviors influenced by the various feelings/emotions. The choice is also affected by the current situation, external and internal conditions, by the relationship between the behaviors, and by the residual activation values of various behaviors. Here in our male, there may have been a previously instantiated goal context hierarchy for avoiding the weasel previously sensed. An appropriate behavior in avoiding the predator may be chosen in spite of the presence of the female vole. Alternatively, an initial step in the behavior stream for approaching and exploring the female vole may win out. 9. Action Taken. The execution of a behavior (goal context) results in the behavior codelets performing their specialized tasks, which may have external or internal consequences, or both. This is IDA taking an action. The acting codelets also include at least one expectation codelet (see Step 6) whose task it is to monitor the action and to try and bring to consciousness any failure in the expected results. If this particular male that has few opportunities to copulate with a female, searching for the female would likely have been selected, resulting in behavior codelets acting to turn the male in the direction of the female, to sniff, and to begin his approach. If on the other hand, our vole has frequent opportunities to mate with females, he may stop his search for this female when he encounters the odor of a weasel or a male conspecific (delBarco-Trillo & Ferkin, 2004). Hypotheses Although our ontology provides a conceptual framework within which to conduct empirical research, its function is not to suggest hypotheses. Formulating hypotheses is one of the functions of mathematical, computational, and conceptual models. Thus, it’s reasonable to formulate potentially testable hypotheses, for the IDA model. By doing so, we hope to stimulate tests of our hypotheses. Here we present a few selected testable hypotheses that may be tested with current technology. 1. The Cognitive Cycle: The very existence of the cognitive cycle in various species, along with its timing (asynchronously cascading at a rate of roughly 5- 10 Hz) is a major hypotheses. Neuroscientists have provided suggestive evidence for this hypothesis (Freeman, 2003; Halgren, Boujon, Clarke, Wang, & Chauvel, 2002; Lehmann, Strik, Henggeler, Koenig, & Koukkou, 1998). Concluding Thoughts The IDA model, constructed within our proposed ontology, represents a theoretical methodology that is radically different from those that appear in the comparative cognition literature. Although the model is heavily based on experimental findings in cognitive and comparative psychology and in brain science, there is only qualitative consistency with experiments. Rather there are a large number of hypotheses (see the preceding section for examples) derived from an unusual computational and conceptual model of cognition, the IDA model. The model is unusual in two significant ways. First, it functionally integrates a broad swath of cognitive phenomena. Second, it does not predict numerical data from experiments but, rather, is implemented as a software agent, IDA. The IDA model generates hypotheses about comparative cognition by way of its design, the mechanisms of its modules, their interaction, and its performance. All of these hypotheses are, in principle, testable. The complex cognitive cycle hypothesized for animals and for humans by the IDA model, samples the world at roughly five to ten times a second. This frequent sampling allows for an exceptionally fine grain analysis of common cognitive phenomena such as a male vole’s reproductive activity (Franklin et al., 2005). At a high level of abstraction these analysis support commonly held explanations of what occurs in these situations and why. At the finer grained level, our analyses flesh out these common explanations, adding detail and functional mechanisms. Therein lies the value of our analyses. Unfortunately, our current techniques for studying comparative cognition at so fine grained a level, that is PET, fMRI, EEG, implanted electrodes, etc., are all lacking either in scope, in spatial resolution, or in temporal resolution. PET and fMRI have temporal resolution problems (Kim, Richter, & Ugurbil, 1997). EEG is well known to have localizability difficulties, and implanted electrodes (in epileptic patients), while excellent in temporal and spatial resolution, can only sample a limited number of neurons, that is, is lacking in scope. As a result, many of our hypotheses, while testable in principle, are not testable at the present time for lack of technologies with suitable scope and resolution. Nonetheless, the integrative nature of the IDA model suggests that these hypotheses will prove useful in helping to guide the research of comparative cognition researchers interested in such cognitive processing. Also, there is the issue of the breadth of the IDA model, encompassing perception, working memory, declarative memory, attention, decision making, procedural learning and more. This breadth raises suspicions. How can such a broad model produce anything useful? The model may not be useful predictor of the processing among animals that do not have the neural circuitry to store such information. For these animals, processing may follow another type of model, which we have not described. Another caveat that we offer is that we have simplified the cognitive and the physiological mechanisms that are likely involved in the processing of such sensory information for our vole. We acknowledge the complexity of physiology and cognition, but the focus of our approach was to provide models that could provide a better understanding of how animals may assess and process information about their social environment. We are confident that researchers in the future will fill in the underlying mechanisms. One of the strengths of the model is that it provides hypotheses, many of which are testable. We believe that the hypotheses suggested in this paper and those that will be developed by other researchers will not only test the predictions of the model, but will also lead to interesting findings and the development of new hypotheses. The IDA model suggests that these various aspects of comparative cognition are so highly integrated in their function that they cannot be fully understood in a highly reductionist manner. A more global view can be expected to add understanding to that produced by the more common, quite specific models. References
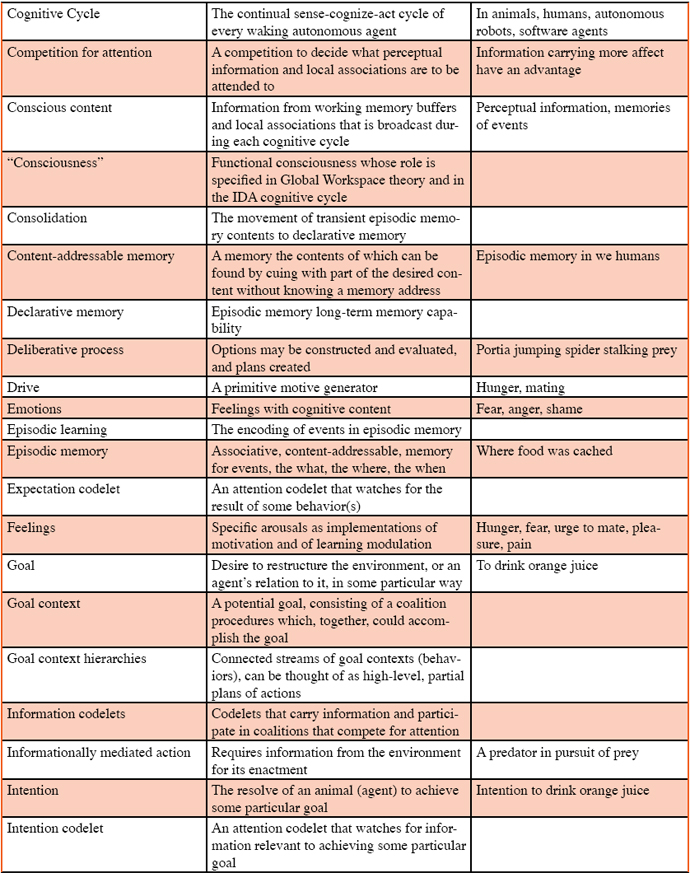
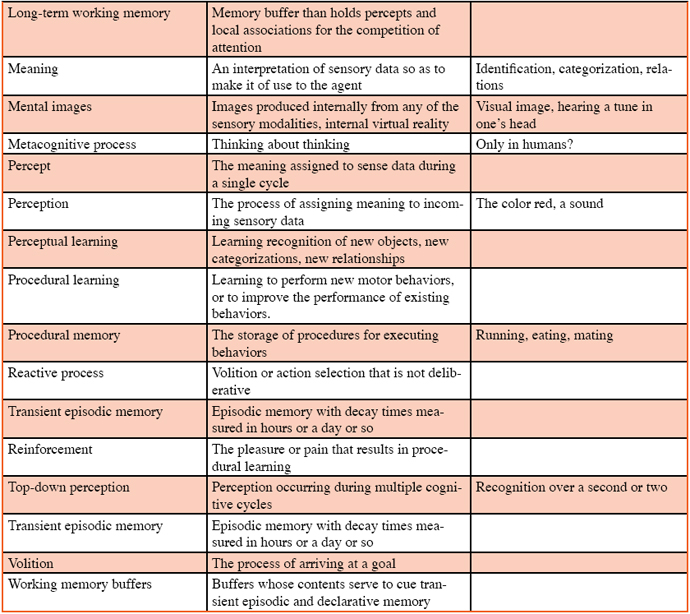
Anderson, M. L. (2003). Embodied cognition: A field guide. Artificial Intelligence, 149, 91-130. Anwar, A., & Franklin, S. (2003). Sparse distributed memory for “conscious” software agents. Cognitive Systems Research, 4, 339–354. Baars, B. J. (1988). A cognitive theory of consciousness. Cambridge: Cambridge University Press. Baars, B. J. (2002). The conscious access hypothesis: Origins and recent evidence. Trends in Cognitive Science, 6, 47–52. Baars, B. J., & Franklin, S. (2003). How conscious experience and working memory interact. Trends in Cognitive Science, 7, 166–172. Baddeley, A. D (1993). Working memory and conscious awareness. In. A. Collins, S. Gathercole, M. A. Conway, & P. Morris (Eds.), Theories of memory. New York: Lawrence Erlbaum Associates. Baddeley, A. D. (2000). The episodic buffer: A new component of working memory? Trends in Cognitive Science, 4, 417–423. Baddeley, A. D., Conway, M. A., & Aggelton, J. (2001). Episodic memory: New directions in research. Oxford: Oxford University Press. Balda, R. P., Pepperberg, I. M., & Kamil, A. C. (1998). Animal cognition in nature: The convergence of psychology and biology in laboratory and field. San Diego: Academic Press. Bekoff, M., Allen, C. & Burghardt, G. M. (2002). The cognitive animal. Cambridge, MA: MIT Press. Blumberg, M. S., & Wasserman, E. A. (1995). Animal mind and the argument from design. American Psychologist 50, 133–144. Bogner, M., Ramamurthy, U., & Franklin, S. (2000). "Consciousness” and conceptual learning in a socially situated agent. In K. Dautenhahn (Ed.), Human cognition and social agent technology (pp.113-135). Amsterdam: John Benjamins. Boysen, S. T., & Himes, G. T. (1999). Current issues and emerging theories in animal cognition. Annual Review of Psychology. 50, 683-705. Brown, S. W., & Bennett, E. D. (2002). The role of practice and automaticity in temporal and nontemporal dual-task performance. Psychological Research, 66, 80-89. Busemeyer, J. R., Medin, D. L., & Hastie, R. (1995). Decision making from the cognitive perspective. The psychology of learning and motivation (vol. 32), Burlington, MA: Academic Press. Clayton, N. S., & Dickinson, A. (1998). Episodic-like memory during cache recovery by scrub jays. Nature, 395, 272-274. Clayton, N. S., Griffiths, D. P., & Dickinson, A. (2001). Elements of episodic-like memory in animals. In A. M. Baddeley, M. A. Conway, & J. Aggelton (Eds.), In Episodic memory: New directions in research (pp. 232-248). Oxford: Oxford University Press. Clayton, N. S., Busey, T. J., & Dickinson, A. (2003). Can animals recall the past and plan for the future? Nature Reviews in Neuroscience, 4, 685-691. Conway, M. A. (2001). Sensory-perceptual episodic memory and its context: Autobiographical memory. In A. Baddeley, M. Conway, & J. Aggleton (Eds.), Episodic memory: New directions in research (pp. 53-70). Oxford: Oxford University Press. Corkin, S. (1968). Acquisition of motor skill after bilateral medial temporal-lobe excision. Neuropsychologia, 6, 255-265. Dalgleish, T., & Power, M. (1999). Handbook of cognition and emotion. Chichester: Wiley. Davis, R., Shrobe, H., & Szolovits, P. (1993). What is a knowledge representation? AI Magazine, 14, 17-33. delBarco-Trillo, J., & Ferkin, M. H. (2004). Male mammals respond to a risk of sperm competition conveyed by odours of conspecific males. Nature, 431, 446-449. Dennett, D. C. (1987). The intentional stance. Cambridge, MA: MIT Press. Edelman, G. M. (1987). Neural darwinism. New York: Basic Books. Eimer, M., & Schlagecken, F. (2003). Response facilitation and inhibition in subliminal priming. Biological Psychology, 64, 7-26. Emery, N. J., & Clayton, N. S. (2001). Effects of experience and social context on prospective caching strategies by scrub jays. Nature, 443-446. Ericsson, K. A., & Kintsch, W. (1995). Long-term working memory. Psychological Review, 102, 211–245. Ferkin, M. H. (1988). The effect of familiarity on social interactions in meadow voles, Microtus pennsylvanicus: A laboratory and field study. Animal Behaviour, 36, 1816-1822. Ferkin, M. H. (1990). Kin recognition and social behavior in microtine rodents. In R. H. Tamarin, R. S. Ostfeld, S. R. Pugh, & G. Bujalska (Eds.), Social systems and population cycles in voles (pp. 11-24). Switzerland: Birkenhaus Press. Ferkin, M. H., & Johnston, R. E. (1995a). Meadow voles, Microtus pennsylvanicus, use multiple sources of scent for sexual recognition. Animal Behaviour ,49, 37-44. Ferkin, M. H., & Johnston, R. E. (1995b). Effects of pregnancy, lactation, and postpartum oestrous on odour signals and the attraction in females meadow voles, Microtus pennsylvanicus. Animal Behaviour, 49, 1211-1217. Ferkin, M. H., Dunsavage, J., & Johnston, R. E. (1999). What kind of information do meadow voles, Microtus pennsylvanicus, use to distinguish between the odors of the top and bottom-scent donors of an over-mark? Journal of Comparative Psychology, 113, 43-51. Ferkin, M. H., Lee, D. N., & Leonard, S. T. (2004a). The reproductive state of female voles affects their scent marking behavior and the responses of male conspecifics to such marks. Ethology, 110, 257-272. Ferkin, M. H., Li, H. Z., & Leonard, S. T. (2004b). Meadow voles and prairie voles differ in the percentage of conspecific marks they over-mark. Acta Ethologica, 7, 1-7. Ferkin, M. H., Mech, S. G., & Paz-y-Mino, C. G. (2001). Scent marking in meadow voles and prairie voles: a test of three hypotheses. Behaviour, 138, 1319-1336. Ferkin, M. H., Tamarin, R. H., & Pugh, S. R. (1992). Cryptic relatedness and the opportunity for kin recognition in microtine rodents. Oikos, 63, 328-332. Ferkin, M. H., Pierce, A. A., Sealand, R. O., & delBarco-Trillo, J. (2005). Meadow voles, Microtus pennsylvanicus, can distinguish more over-marks from fewer over-marks. Animal Cognition, 8, 82-89. Franklin, S. (1995). Artificial minds. Cambridge, MA: MIT Press. Franklin, S. (2000). Deliberation and voluntary action in ‘conscious’ software agents. Neural Network World, 10, 505–521. Franklin, S. (2001a.) Automating human information agents. In Z. Chen & L. C. Jain (Eds.), Practical applications of intelligent agents (pp. 27-58). Berlin: Springer-Verlag. Franklin, S. (2001b). Conscious software: A computational view of mind. In V. Loia & S. Sessa (Eds.), Soft computing agents: New trends for designing autonomous systems (pp. 1-46). Berlin: Springer: Physica-Verlag. Franklin, S. (2005a). A “consciousness” based architecture for a functioning mind. In D. Davis (Ed.), Visions of mind (pp. 149-175). Hershey, PA: IDEA Group, Inc. Franklin, S. (2005b). Evolutionary pressures and a stable world for animals and robots: A commentary on Merker. Consciousness and Cognition, 14, 115-118. Franklin, S. (2005c). Perceptual memory and learning: Recognizing, categorizing, and relating. Symposium on developmental robotics (pp. 1-46), American Association for Artifical Intelligence (AAAI). Stanford University, Palo Alto, CA, USA. March 21-23, 2005. Franklin, S., Baars, B. J., Ramamurthy, U., & Ventura, M. (2005). The role of consciousness in memory. Brains, Minds and Media, 1, 1-38. Franklin, S., & Graesser, A. C. (1997). Is it an agent, or just a program?: A taxonomy for autonomous agents. In J. P. Mueller, M. J. Wooldridge, & N. R. Jennings (Eds.), Intelligent Agents III (pp. 21-35). Berlin: Springer Verlag. Franklin, S., & McCauley, L (2004). Feelings and emotions as motivators and learning facilitators. Architectures for modeling emotion: Cross-disciplinary foundations, AAAI 2004 Spring Symposium Series; American Association for Artificial Intelligence. Stanford University, Palo Alto, CA, USA. March 22-24, 2004. Freeman, W. J. (1999). How brains make up their minds. London: Weidenfeld & Nicolson General. Freeman, W. J. (2003). Evidence from human scalp electroencephalograms of global chaotic itinerancy. Chaos, 13, 1067-1077. Glenberg, A. M. (1997). What memory is for. Behavioral and Brain Sciences, 20, 1-19. Griffin, D. R. (1976). The question of animal awareness: Evolutionary continuity of mental experience. New York: Rockefeller Press. Griffin, D. R. (1984). Animal thinking. Cambridge: Harvard University Press. Griffin, D. R. (2001). Animal minds: Beyond cognition to consciousness (2nd edition). Chicago: University of Chicago Press. Griffiths, D., Dickinson, A., & Clayton, N. S. (1999). Episodic memory: What can animals remember about their past? Trends in Cognitive Science, 3, 74-80. Hacker, D. J., & Bol, L. (2004). Metcognitive theory: Considering the social-cognitive influences. In D. M. McInerney & S. Van Etten (Eds.), Big theories revisited (pp. 275-297). Greenwich, CT: Information Age Press. Halgren, E., Boujon, C., Clarke, J., Wang, C., & Chauvel, P. (2002). Rapid distributed fronto-parieto-occipital processing stages during working memory in humans. Cerebral Cortex, 12, 710-728. Harnad, S. (1990). The symbol grounding problem. Physica D, 42, 335-346. Hauser, M. D. (2000). Wild animals: What animals really think. New York: Henry Holt. Hirsh, I. J. (1996). Auditory psychophsyics and perception. Annual Review of Psychology, 47, 461-484. Hofstadter, D. R., & Mitchell, M. (1995). The copycat project: A model of mental fluidity and analogy-making. In K. J. Holyoak & J. A. Barnden (Eds.), Advances in connectionist and neural computation theory Vol. 2: Logical connections. Norwood, NJ: Ablex. Hulse, S., Fowler, H, & Honig, W. (1978). Cognitive processes in animal behavior. Mahwah, NJ: Lawrence Erlbaum Associates. Jackson, J. V. (1987). Idea for a mind. Siggart Newsletter, 181, 23–26. Kamil, A. C. (1998). On the proper definition of cognitive ethology. In R. P. Balda, I. M. Pepperberg & A. C. Kamil (Eds.), Animal cognition in nature: The convergence of psychology and biology in laboratory and field (pp. 1-28). A. New York: Academic Press. Kahneman, D., Slovic, P., & Tversky, A. (1982). Judgement under uncertainty: Heuristics and biases. Cambridge: Cambridge University Press. Kanerva, P. (1988). Sparse distributed memory. Cambridge, MA: MIT Press. Kim, S.-G., Richter, W., & Ugurbil, K. (1997). Limitations of temporal resolution in functional MRI. Magnetic Resonance Medicine, 37, 631-636. Koch, C. (2004). The Quest for Consciousness: A neurobiological approach. Englewood, Colorado: Roberts & Co. Kuhn, T. (1970). The structure of scientific revolutions (2nd ed.). Chicago: University of Chicago Press. Lakatos, I. (1974). The methodology of scientific research programs. Cambridge: Cambridge University Press. Langston, C. (1989). Artificial life. Redwood City, CA: Addison-Wesley. Lehmann, D., Strik, W. K., Henggeler, B., Koenig, T., & Koukkou, M. (1998). Brain electric microstates and momentary conscious mind states as building blocks of spontaneous thinking: I. Visual imagery and abstract thoughts. International Journal of Psychophysiology, 29, 1-11. Lisman, J. E., & Fallon, J. R. (1999). What maintains memories? Science, 283, 339-340. Lu, Z., & Sperlin, G. (2001). Three-systems theory of human visual motion perception: Review and update. Journal of Optical Society of America. A 18, 2331-2370. Maes, P. (1989). How to do the right thing. Connection Science, 1, 291–323. Maturana, R. H., & Varela, F. J. (1980). Autopoiesis and cognition: The realization of the living. Dordrecht, Netherlands: Reidel. Maturana, H. R. (1975). The organization of the living: A theory of the living organization. International Journal of Man-Machine Studies, 7, 313–332. Merker, B. (2005). The liabilities of mobility: A selection pressure for the transition to consciousness in animal evolution. Consciousness and Cognition, 14, 89-114. Mounoud, P. 1993. The emergence of new skills: Dialectic relations between knowledge systems. In G. J. P. Savelsbergh (Ed.), The development of coordination in infancy (pp. 13-46). Amsterdam: North Holland. Nadel, L. (1992). Multiple memory systems: What and why. Journal of Cognitive Neuroscience, 4, 179-188. Negatu, A., & Franklin, S. (2002). An action selection mechanism for ‘conscious’ software agents. Cognitive Science Quarterly, 2, 363–386. O’Connor, K. P, & Aardema, F. (2005). The imagination: Cognitive, pre-cognitive, and meta-cognitive aspects. Consciousness and Cognition, 14, 233-256. O’Craven, K., & Kanwisher, N. (2000). Mental imagery of faces and places activates corresponding stimulus-specific brain regions. Journal of Cognitive Neuroscience, 12, 1013–1023. Olton, D. S., & Samuelson, R. J. (1976). Remembrance of places passed: spatial memory in rats. Journal of Experimental Psychology: Animal Behavior Processes, 2, 97–116. Oyama, S. (1985). The ontogeny of information. Cambridge: Cambridge University Press. Panksepp, J. (1998). Affective neuroscience: The foundations of human and animal emotions. Oxford: Oxford University Press. Papaj, D. R., & Lewis, A. C. (1993). Insect learning: Ecological and evolutionary perspectives. New York: Chapman & Hall. Perr, R. J., & Hodges, J. R. (1999). Attention and executive deficits in Alzheimer’s disease. Brain, 122, 383-404. Proctor, R., Wang, R. H., & Vu, K-P., L. (2002). Influences of conceptual, physical, and structural similarity on stimulus-response compatibility. Quarterly Journal of Experimental Psychology, 55A, 59-74. Pylyshyn, Z. (1999). Is vision continuous with cognition? The case for cognitive impenetrability of visual perception. Behavioral and Brain Sciences, 22, 341-365. Rauchs, G., Desgranges, B., Foret, J., & Eustache, F. (2005). The relationships between memory systems and sleep stages. Journal of Sleep Research, 14, 123-140. Reber, A. S., Walkenfeld, F. F., & Hernstadt, R. (1991). Implicit and explicit learning: Individual differences and IQ. Journal of Experimental Psychology: Learning, Memory, and Cognition, 17, 888-896. Roskies, A. L. (1999). The binding problem. Neuron, 24, 7-9. Schacter, D. L., & Tulving, E. (1994). Memory systems. Cambridge, MA: MIT Press. Schmidt, R. A. (1988). Motor control and learning: A behavioral emphasis (2nd ed). Champaign, IL: Human Kinetics. Searle, J. R. (1983). Intentionality: An essay in the philosophy of mind. Cambridge: Cambridge University Press. Shanahan, M. P. (2005.) Consciousness, emotion, and imagination: A brain-inspired architecture for cognitive robotics. Proceedings AISB 2005 Symposium on Next Generation Approaches to Machine Consciousness. Shettleworth, S. J. (1998). Cognition, evolution, and behavior. Oxford: Oxford University Press. Sloman, A. (1987). Motives mechanisms emotions. Cognition, 1, 217–234. Sloman, A. (1999). What sort of architecture is required for a human-like agent? In. M. Wooldridge & A. S. Rao (Eds.), Foundations of Rational Agency (pp. 35-52). Dordrecht, Netherlands: Kluwer Academic Publishers. Smith, C., & MacNeill, C. (1994). Impaired motor memory for a pursuit rotor task following Stage 2 sleep loss in college students. Journal of Sleep Research, 3, 206-213. Smith, J. D., & Washburn, D. A. (2005). Uncertainty monitoring and metacognition by animals. Current Directions in Psychological Science, 14, 19-24. Standing, L. (1973). Learning 10,000 pictures. Quarterly Journal of Experimental Psychology, 25, 207-222. van Gorp, W. Altshuler, G. L., Theberge, D. C., & Mintz, J. (1999). Declarative and procedural memory in bipolar disorder. Biological Psychiatry, 46, 525-531. Wasserman, E. A. (1993). Comparative cognition: Beginning the second century of the study of animal intelligence. Psychological Bulletin, 113, 211–228. Wilcox, S., & Jackson, R. (1998). Cognitive abilities of araneophagic jumping spiders tricksters: Deceit, predation, and cognition. In R. P. Balda, I. M. Pepperberg, & A. C. Kamil (Eds.), Animal cognition in nature: The convergence of psychology and biology in laboratory and field (pp. 411-434). New York: Academic Press. Wulf, G., Hoss, M. & Prinz, W. (2001). Directing attention to movement effects enhances learning. Psychonomic Bulletin & Review, 8, 648-660. Zhang, Z., Franklin, S., Olde, B., Wan, Y., & Graesser, A. (1998). Natural language sensing for autonomous agents. Proceedings of IEEE International Joint Symposia on Intelligence Systems 98. Glossary Table
|
S. Franklin’s work on the IDA model was supported by ONR. M. H. Ferkin was supported by grants from the NIH AG16594- 01 and MH61971 and NSF IBN 9421529 and IOB 0444553. We are indebted to B.J. Baars and to members of the “Conscious” Software Research Group. We thank the editors and anonymous reviewers for their many helpful suggestions. |