| <<Vol. 2 Table of Contents | |
|
|
|
|
2007 |
Volume 2, pp 47-66 |

Comparative Cognition, Hippocampal Function, and Recollection
Howard Eichenbaum
Boston University
|
|
||||||||||||||||||||||||
|
||||||||||||||||||||||||
|
Aristotle (350BC) contended that "…other animals (as well as man) have memory, but, of all that we are acquainted with, none, we venture to say, except man, shares in the faculty of recollection." Here Aristotle distinguishes "memory," as an elementary matching of current sensations to impressions from prior experience, from the capacity for reminiscence, as the ability to mentally reconstruct past experiences. Jumping to current times, Tulving (1983, 2002) characterized animals as having the capacity for acquiring semantic knowledge, but claimed that episodic recollection "… has evolved only once, and in only one species, although other species would benefit from it as much as do humans." Neither Aristotle nor Tulving employed experimental evidence in support of their claims, but several other contemporary researchers have, with indecisive results, such that there is currently no consensus on whether animals have the capacity for recollection (reviewed in Clayton, Bussey, & Dickinson, 2003; Clayton, Bussey, Emery, & Dickinson, 2003; Hampson & Schwartz, 2004; see also Roberts, 2005). This literature will not be critiqued again here. Instead, I will argue simply that the issue of recollection in animals can be addressed. Those who define recollection as accessible to consciousness and available to declarative or explicit expression will not find solace in this review. However, those who define recollection in terms of the organization of its contents have generated a vigorous area of study that is addressing the question. Here I will present one perspective on that approach. First I will consider how we might define recollection in terms of features of memory that can be assessed in animals as well as humans, and in doing so introduce three fundamental features of recollection that can be examined across species. I will then introduce the cortical-hippocampal system that supports recollection in humans, focusing on the role of the hippocampus. I will consider evidence, using examples primarily from experiments in my own laboratory, indicating that animals exhibit each of the defining features of recollection, that the hippocampus is critical to these features of recollection in animals, and that neurons in the hippocampus encode information that support these features of recollection. I will briefly discuss the often emphasized role of the hippocampus in spatial function in animals, offering a view on how the data on spatial navigation and memory can be reconciled with a more general role for the hippocampus in recollection. Finally, I will consider the anatomical organization of the hippocampus and associated cortical areas, and the functions of these areas, in several species. The following considerations are based on recent and more extensive reviews covering each of these issues (Eichenbaum, 2004; Eichenbaum, Fortin, Ergorul, Wright, & Agster, 2005; Manns & Eichenbaum, 2005, 2006; Eichenbaum, Yonelinas, & Ranganath, 2006), combined here to address the question of whether recollection is a cognitive function that is conserved across mammalian species. Importantly, I will not present data from a large number of animal species, as might be expected by some in a comprehensive comparative review. Instead I will focus on a few highly domesticated species of rodents, and a few non-human primate species that have been the subject of extensive behavioral and neurobiological investigations, and I make no apology for this focus. My aim is to identify fundamental features of recollection that can, at least in principle, be studied in any species and to identify the brain circuitry that supports common features of recollection in mammals. Indeed, because there is an extensive literature on these issues in laboratory rodents and monkeys, one can make conclusions that are generalizable to a variety of tests of memory function and different kinds of investigations on the relevant brain areas. I would predict that the same features of recollection and analogous roles of the same brain areas can be identified in less studied, non-domesticated species and that the interpretations offered here will be relevant to the natural use of memory in their specialized habitats. The Brain System that Supports Recollection The brain system that supports recollection involves a network of widespread cortical association areas and structures in the medial temporal lobe (Eichenbaum, 1999). The cortical areas involve components of the prefrontal cortex, as well as structures of the diencephalon, that mediate working memory, effortful retrieval, source monitoring, and other processing that contribute critically to cognitive functions essential to recollection (e.g., Aggleton & Brown, 2006; Henson, Shallice, Josephs, & Dolan, 1999; Yonelinas, Otten, Shaw, & Rugg, 2005). Also, areas of the parietal and temporal cortex are involved in complex perceptual processing essential to configuration of the conceptual contents of information that is the subject of recollection (e.g., Uncapher, Otten, & Rugg, 2006). Projections from these areas strongly converge onto the medial temporal lobe, which also sends strong projections back to these cortical areas, suggesting a central role in organizing or extending the persistence of cortical representations. Damage to the medial temporal area, including the hippocampus and surrounding parahippocampal cortical areas, results in a profound deficit in encoding information in a way that is subsequently subject to recollection (for review, see Eichenbaum & Cohen, 2001). Furthermore, unlike the cortical areas of this system, the role of the medial temporal lobe is fully selective to memory. Therefore, this review will focus specifically on the components of the medial temporal lobe, and in particular on the hippocampus. What is Recollection? I will begin the discussion by considering the distinction between a vivid recollection and something less, a sense of familiarity with a particular person or object. We have all been in the situation where we meet someone who seems slightly or perhaps highly familiar but we cannot recall who they are or why we know them. Sometimes, we just give up and say, "Don’t I know you?". Alternatively, that embarrassment is sometimes avoided when a clue or sufficient mental searching helps us suddenly retrieve the name, where we met before, and the circumstances of the meeting. Familiarity is rapid and defined in terms of the strength of the match of a cue to a stored memory template. It is an isolated ability to identify a current stimulus (a person or object) as previously experienced. Recollection is typically slower and is defined by the number of associations retrieved and the organization of the memory obtained. Thus, recollections typically include not only the item sought in memory but also the spatial and temporal context of the experience in which the item was previously encountered. Furthermore, our most vivid recollections involve replaying an entire episode in which we met the person, and that memory might lead to remembering additional encounters. These considerations tell us that familiarity and recollection differ both in the dynamics of memory retrieval and in the contents of what is retrieved. These properties of recollection will be the subject of the comparative analysis presented here. The Role of the Hippocampus in Recollection and Familiarity As the incident described above suggests, one of the ways familiarity and recollection are distinguished is by their retrieval dynamics. Familiarity occurs quickly and is graded in strength. Items from our past can generate a slight sense of familiarity or an intensely held belief that we have experienced them before. By contrast, recollection is qualitative. Its goodness is characterized by the number of associations we retrieve, and we tend to retrieve each one in an all-or-none fashion. How can these properties be dissociated in the performance of human and animal subjects? The retrieval dynamics of recollection and familiarity have been distinguished in humans by the analysis of receiver operating characteristic (ROC) functions derived from recognition memory performance (Yonelinas, 2001). In a typical experiment, subjects study a list of words, then are tested for their capacity to identify as "old" or "new" the same words plus a set of words that were not studied. The resulting ROC analysis plots "hits," that is, correct identifications of old items, against "false alarms," incorrect identifications of new items as if they were old, across a range of confidence levels. This analysis typically reveals an asymmetric function characterized by an above-zero threshold of recognition at the most conservative criterion (zero false alarm rate) and thereafter a curvilinear performance function (Yonelinas, 2001; Figure 1a). The positive Y-intercept is viewed as an index of the recollection in the absence of measurable familiarity, whereas the degree of curvature reflects familiarity as typical of a signal-detection process (Macmillan & Creelman, 1991). Consistent with this view, under different experimental demands that favor one of these processes, the shape of the ROC curve takes on distinguishable functions (Yonelinas, 2001). During performance that favors recollection, the ROC curve highlights the threshold component of recognition with performance at successively higher confidence levels characterized by a linear function (Figure 1b). In contrast, during performance that favors familiarity, the ROC curve is symmetrical and curvilinear (Figure 1c).
Yonelinas et al. (2002) used ROC analysis to show that mild hypoxia that causes damage largely confined to the hippocampus resulted in a severe deficit in recollection but normal familiarity. The distinction between impaired recollection and spared familiarity was verified by measures of subjective experiences in recognition reflected in "remember" versus "know" judgments by the same patients. In addition, structural equation modeling methods used on a large sample of hypoxic patients revealed that hypoxic severity predicted the degree to which recollection, but not familiarity, was impaired. A similar pattern of deficient recollection and preserved familiarity was reported in a patient with relatively selective hippocampal atrophy related to meningitis (Aggleton et al., 2005). These studies implicate the hippocampus as playing a selective role in recollection. However, other interpretations of the data on ROC analyses in normal human subjects have led to the view that recollection and familiarity reflect differences in strength of a single memory function (Wixted & Stretch, 2004), and indeed many reports are mixed on whether ROC curves are more consistent with single or dual processes in recognition, suggesting that the dissociation of these processing functions may be dependent on parameters of testing and assumptions in the data analysis. In addition, another ROC study reported deficits in both recollection and familiarity in hypoxic patients with identified hippocampal damage (Wais, Wixted, Hopkins, & Squire, 2006), and several other studies also reflect a mixture of results on whether the hippocampus is selectively involved in recollection or involved in both recollection and familiarity. Differences in the localization of damage in different patients as well as differences in the task demands across studies might account for the variability in results across these studies. To address whether recollection and familiarity can be distinguished in ROC functions by selective hippocampal damage, we developed an ROC protocol for assessing recollection and familiarity in rats and for examining the effects of highly selective experimental lesions of the hippocampus. Our recognition task exploited rats’ superb memory capacities with odors (Fortin, Wright, & Eichenbaum, 2004). On each daily test session, rats initially sampled 10 common household scents mixed in with playground sand in a plastic cup containing a cereal reward (Video 1). When each sample was presented the animal would dig for the reward and incidentally smell the odor of the sand. Following a 30 minute memory delay, the same odors plus 10 additional odors were presented one at a time in random order. On each recognition test, the animal followed a non-match to sample rule such that it could dig in the target odor to obtain a reward if the target was "new" (a non-match) or could refrain from digging if the odor was "old" (a match) and instead obtain a reward in an empty cup on the opposite end of the test chamber. Initially animals were trained with short lists of odors, and the list length was gradually increased to 10 items. In addition, in the final phase of training and testing, a different response criterion was encouraged for each daily session using a combination of variations in the height of the test cup, making it more or less difficult to respond to that cup, and manipulations of the reward magnitudes associated with correct responses to the test and the unscented cup. Notably, the use of a method for explicitly varying the animal’s bias is different from the use of confidence judgments in experiments on recognition in humans (Yonelinas, 2001); nevertheless, both methods successfully vary the subject’s criterion along the full range required to compute ROC curves.
The ROC curve of intact control rats was asymmetric (Figure 2), containing both a threshold component (above-zero Y-intercept) and a strong curvilinear component. This pattern is remarkably similar to the ROC of humans in verbal recognition performance (Figure 1a), consistent with a combination of recollection-like and familiarity-based components of recognition in animals. To explore the role of the hippocampus in recollection, subjects were subsequently divided into two groups matched on both performance components, and one group received selective lesions of the hippocampus whereas the other group received sham control operations. After recovery, we again tested recognition performance at each response criterion. The ROC of control rats continued to reflect both recollection-like and familiarity components, whereas the ROC of animals with selective hippocampal lesions was fully symmetrical and curvilinear (Figure 2), characteristic of familiarity-based recognition in humans (Figure 1c). To describe these patterns quantitatively, we calculated indices of recollection and familiarity (Figure 2 insets). Whereas familiarity remains normal in rats with hippocampal lesions, recollection is severely impaired. The overall level of performance (averaged across biases) on the task is slightly worse in the hippocampal group (66%, compared to 73% in controls). Given that any performance deficit would be expected to result in an ROC closer to the diagonal (chance performance; dashed line in Figure 2), it is possible that the alteration in their ROC pattern resulting from the hippocampal lesion reflect a generalized decline in memory. In order to compare the ROC of hippocampal rats with the pattern of forgetting in normal animals, we challenged the memory of control rats by increasing the memory delay to 75 minutes. This manipulation succeeded in reducing the overall level of performance of control animals to 64%, equivalent to that of the hippocampal rats. Yet, further testing of the controls showed that their ROC continued to have an asymmetrical threshold component, as indicated by an above-zero Y-intercept. Notably, the controls’ ROC was distinctly more linear than that of both the hippocampal rats and the controls when tested at the shorter memory delay. This pattern of performance suggests that, in normal rats, familiarity fades more quickly than recollection, similar to observations on humans (Yonelinas, 2002). Moreover, comparison of the ROC curve in normal rats at the 75 minute delay versus that of rats with hippocampal damage at the 30 minute delay emphasizes the distinction between these two groups in their differential emphasis on recollection and familiarity, respectively, even when the overall levels of recognition success are equivalent. These findings strongly suggest that rats exhibit two distinct processes in recognition, one that is marked by a threshold retrieval dynamic characteristic of episodic recollection in humans, and another that follows a symmetrical and curvilinear processing function characteristic of familiarity in humans. These observations suggest comparable dual retrieval mechanisms underlying recognition in animals and humans, and strongly support the notion that the hippocampus plays a critical role only in the recollective processes that contribute to recognition. Fundamental Features of the Contents of Recollection The above described experiment provides evidence suggesting the retrieval dynamics of recollection are similar in animals and humans. Further evidence suggesting conservation of recollective function across species can be found in an examination of the contents of recollected memories. Recollection in humans is highlighted by three central features of its contents. First, when we move beyond a sense of familiarity with a previously experienced stimulus, we recover information about the context or source in which the stimulus was experienced, most typically "where" and "when" an event occurred. This aspect of recollection has been investigated in many studies of recollection in humans, as well as animals. Second, vivid recollection is also typically characterized by a "replay" of the sequence of events that occurred in an experience. The capacity for mental replay has been highlighted in Tulving’s (1983) account of episodic memory. Third, recall of one memory often leads to the recollection of a larger set of related memories. This indicates that recollection typically accesses not just one isolated memory, but rather a network of memories. Furthermore, these memories’ networks can be used to infer indirect and novel relationships between elements of those memories, and we employ these insights in a variety of creative ways. Do animals have the capacity for all of these hallmark features of recollection? Which of these features of recollection depends on the hippocampus? In the following sections I will outline experimental studies we have pursued to address these questions. Memory for Where and When Events Occurred Several investigators have argued that animals are indeed capable of remembering the spatial and temporal context in which they experienced specific stimuli (Clayton, Bussey, & Dickinson, 2003; Day, Langston, & Morris, 2003). To further explore these aspects of episodic memory, we developed a task that assesses memory for events from a series of events that each involve the combination of an odor ("what"), the place in which it was experienced ("where"), and the order in which the presentations occurred ("when"; Ergorul & Eichenbaum, 2004). On each of a series of events, rats sampled an odor in a unique place along the periphery of a large open field (Figure 3a; Video 2a). Then, memory for the when those events occurred was tested by presenting a choice between an arbitrarily selected pair of the odor cups in their original locations (Video 2b). We identified the stimulus initially approached and distinguished that response from the final choice in which the rat dug for food. Over a series of shaping phases, rats were trained to select the earlier presented odor of a pair randomly selected from the series.
Rats performed well above chance (76.2% correct) in their choices on the test phase, indicating that they can remember the order of unique sequences of odors and places (Figure 3b). In addition, we also found that rats first approached the correct stimulus at well above chance level, indicating they remembered the sequence of places where the cups were presented prior to perceiving information about the odor at that location; importantly, separate tests showed that rats cannot accurately judge the odor in a cup until they arrive within at the edge of the cup. However, performance was not as accurate in the first approach as it was in the final choice, suggesting that rats begin by guessing the location of the earlier experienced cup, then confirm this choice using the smell of the cup. This hypothesis was confirmed in a control condition in which we presented the test cups without odors. In this condition, performance of intact animals fell to chance, indicating that when the selected location is not confirmed by the associated odor, performance is disrupted (Figure 3b). This pattern of results strongly suggests rats normally use a combination of "where" and "what" information to judge "when" the events occurred. To examine the role of the hippocampus, animals were subsequently separated into matched groups, one of which received selective hippocampal lesions. Subsequently, intact rats continued to choose well on the standard "what-where-when" trials (Figure 3c). By contrast, the performance of animals with hippocampal lesions was no better than chance. In addition, whereas intact rats continued to perform well on the initial approach, rats with hippocampal lesions approached the correct choice less often than expected by chance. Contrary to the strategy of normal rats and the reinforcement contingency of the test phase, rats with hippocampal damage were inclined to visit the more recently presented and rewarded place rather than the earlier visited locus. This observation indicates an intact spatial memory in rats with hippocampal damage, and this memory was employed despite its maladaptive consequences. These findings indicate that the hippocampus is critical for effectively combining the "what," "when," and "where" qualities of each experience to compose the retrieved memory. Normal rats initially employ their memory of the places of presented cups and approach the location of the earlier experience. Then they confirm the presence of the correct odor in that location. Animals with hippocampal damage fail on both aspects of this task and, instead, their behavior is guided by another form of memory that leads to the incorrect first approach. That they can initially approach the most recently rewarded location indicated their spatial memory is intact. However, it appears they are driven to approach the last rewarded cup rather than combine the what-where-when cues to select the earlier event. Memory for the Order of Events Within a Unique Experience In addition to memory for the spatial and temporal context of distinct events, a vivid recollection often involves recalling the flow of events within a single experience. To investigate the memory for the order of events in a unique experience, we developed a behavioral protocol that assesses memory for episodes composed of a unique sequence of olfactory stimuli presented to the animal as it remains in its cage (Fortin, Agster, & Eichenbaum, 2002; see also Kesner, Gilbert, & Barua, 2002). In addition, our design allowed us to directly compare memory for the sequential order of odor events with recognition of the odors in the list independent of memory for their order. On each trial, rats were presented with a series of five odors, selected randomly from a large pool of common household scents. Memory for each series was subsequently probed by a choice test where the animal was reinforced for selecting the earlier of two of the odors that had appeared in the series. For example, the rat might be initially presented with odors A then B then C then D then E. Following the delay, two non-adjacent odors, e.g., B and D, were presented, and the animal would be rewarded for selecting odor that appeared earlier (in this case, B). On each trial, any pair of non-adjacent odors might be presented as the probe, so the animal had to remember the entire sequence in order to perform well throughout the testing session.
After training over many days, rats performed sequential order judgments well above chance levels (Figure 4), indicating they can remember the order of a sequence of events in unique experiences. In order to examine the role of the hippocampus in memory for the order of events in unique experiences, these subjects were divided into two groups matched for performance, and animals in one group were given selective hippocampal lesions whereas those in the other group received sham operations. After recovery, all animals were tested again on memory for the order of odors in unique odor sequences. Intact rats continued to perform well whereas rats with hippocampal lesions were severely impaired, performing no better than chance except when the judgment was easiest (when the odors were first and last in the series). The same rats were then also tested on their ability to recognize odors that were presented in the series. On each trial, a series of five odors was presented in a format identical to that used in the previous testing. Then, recognition was probed using a choice test in which the animal was presented with one of the odors from the series and another odor from the pool that was not in the series (in which food was buried). For example, the rat might instead be presented with the series A through E, then, following a delay, an odor selected randomly from those initially sampled and an odor not presented in the sequence, e.g., A and X, were presented. The rat would be rewarded for choosing X. Both intact rats and rats with selective hippocampal damage acquired the task rapidly, and there was no overall performance difference between the groups in acquisition rate or final level of recognition performance (Figure 4). Furthermore, in both groups, recognition scores were consistently superior on probes involving odors that appeared later in the series, suggesting some forgetting of items that had to be remembered for a longer period and through more intervening items. A potential confound in any study that employs time as a critical dimension in episodic memory is that memories obtained at different times are likely to differ in the strength of their memory traces, due to the inherent decremental nature of memory traces. To what extent could normal animals be using differences in the relative strengths of memory traces for the odors to judge their sequential order? The observation of a temporal gradient in recognition performance by normal animals suggests that memories were in fact stronger for the more recently presented items in each sequence. These differences in trace strength potentially provide sufficient signals for the animals to judge the order of their presentation. However, the observation of the same temporal gradient of recognition performance in rats with hippocampal damage indicated that they had normal access to the differences in trace strengths for the odors. Yet these intact trace-strength differences were not sufficient to support above chance performance in the order probes. These considerations strongly suggest that normal rats also could not utilize the relative strengths of memories for the recently experienced odors, and instead based their sequential order judgments directly on remembering the odor sequence. The findings indicate that animals have the capacity to recollect the flow of events in unique experiences and that the hippocampus plays a critical role in this capacity. Networking Memories A third defining quality of recollection is our capacity to bring to mind multiple related memories, that is, memories that have common elements, and to make inferences from the information contained in those memories. In order to examine the extent to which animals can link memories that share common elements, we studied whether rats can learn a set of odor problems that share elements, and then tested whether they had integrated the memories into networks that support inferential judgments. One experiment compared the ability of normal rats and rats with selective damage to the hippocampus on their ability to learn a set of paired associate problems that contained common elements and to interleave the representations of these problems in support of novel inferential judgments (Bunsey & Eichenbaum, 1996). Animals were initially trained on two sets of overlapping odor paired associates (e.g., A goes with B, B goes with C). Then the rats were given probe tests to determine if they could infer the relationships between items that were only indirectly associated through the common elements (e.g., A goes with C?). Normal rats learned the paired associates and showed strong transitivity in the probe tests (Figure 5). Rats with selective hippocampal lesions also learned the pairs over several trials but were severely impaired in the probes, showing no evidence of transitivity.
In another experiment, rats learned a hierarchical series of premises that involved odor choice judgments between overlapping elements (e.g., A > B, B > C, C > D, D > E), then were probed on the relationship between indirectly related items (e.g., B vs. D?; Figure 6). Normal rats learned the series and showed robust transitive inference on the probe tests. Rats with hippocampal damage also learned each of the initial premises but failed to show transitivity (Dusek & Eichenbaum, 1997). The combined findings from these studies show that rats with hippocampal damage can learn even complex associations, such as those embodied in the odor paired-associates and conditional discriminations. However, without a hippocampus, they do not interleave the distinct experiences by their common elements to form a relational network that supports inferential memory expression. Importantly, according to the present view, the hippocampus does not compute or directly mediate transitive judgments. Rather, the hippocampus supports the encoding and retrieval of information about previous experiences on which cortical areas might accomplish the critical judgment. One neocortical association area that receives hippocampal outputs and is likely critical to inferential judgments is the prefrontal cortex (Waltz et al., 1999).
Neural Representations that Support Recollection Additional insights about the fundamental properties of memory representation can be gained through the analysis of neural activity patterns associated with the critical stimuli and behavioral events that occur in animals performing memory tasks. These studies can confirm the evidence from tests of brain damage by providing evidence of normal coding of features of memory that are lost following selective damage of the same brain areas. In addition, these studies can provide insights about where and how particular types of information are encoded within the circuitry of the hippocampus and associated brain structures. Observations from rats, monkeys, and humans, accumulated across many different behavioral protocols, show that hippocampal neuronal activity reflects each of the three fundamental features of recollection discussed above: representation of events as items in the context in which they are experienced, representation of episodes as sequences of events, and representation of common features of experiences that link distinct memories into networks. Events are represented as items in context A wealth of studies have shown that hippocampal neurons fire associated with the ongoing behavior and the context of events as well as the animal’s location (Eichenbaum, 2004). The combination of spatial and non-spatial features of events captured by hippocampal neuronal activity is consistent with the view that the hippocampus encodes many features of events and the places where they occur. Two recent studies provide examples that highlight the rapid associative coding of events and places by hippocampal neurons. In one study rats were trained on an auditory fear conditioning task in which a tone was paired with shock to produce conditioned freezing to subsequent tone presentations (Moita, Moisis, Zhou, LeDoux, & Blair, 2003). Prior to fear conditioning, few hippocampal cells were activated by the auditory stimulus. Following pairings of tone presentations and shocks, many cells fired briskly to the tone and did so only when the animal was in a particular place where the cell had fired above baseline prior to conditioning. Another study examined the firing properties of hippocampal neurons in monkeys performing a task where they rapidly learned new scene-location associations (Wirth et al., 2003). Just as the monkeys acquired a new response to a location in the scene, neurons in the hippocampus changed their firing patterns to become selective to particular scenes.
Additional studies have directly examined the extent to which hippocampal neurons encode specific stimuli and places where they occur by training subjects to perform the same memory judgments at many locations in the environment. In one study, rats performed a task in which they had to recognize any of nine olfactory cues when placed in any of nine locations (Wood, Dudchenko, & Eichenbaum, 1999; Figure 7). On each trial, the rat was rewarded when it responded to a cue that differed from (was a non-match to) the immediately preceding stimulus. Because the location of the discriminative stimuli was varied systematically, cellular activity related to the stimuli and behavior could be dissociated from that related to the animal’s location. Some hippocampal cells encoded particular odor stimuli, others were activated when the rat sampled any odor at a particular place, and yet others fired associated with whether the odor matched or differed from the previous cue (Figure 8). However, the largest subset of hippocampal neurons fired only associated with a particular combination of the odor, the place where it was sampled, and the match-non-match status of the odor. In a similar task created for humans, Ekstrom et al. (2003) recorded the activity of hippocampal neurons as people played a taxi driver game, searching for passengers picked up and dropped off at various locations in a virtual reality town. Some cells encoded particular cues or fired as the subject traversed specific locations. Also, many of these cells fired selectively when the subject viewed a particular scene from a particular place or passed a location while pursuing a particular goal. Hippocampal cells that represent specific salient objects in the context of a particular environment have also been observed in studies of rats engaged in foraging (Gothard, Skaggs, Moore, & McNaughton, 1996; Rivard, Lenck-Santini, Poucet, & Muller, 2004) and place learning (Hollup, Molden, Donnett, Moser, & Moser, 2001) in open fields. Furthermore, parallel evidence from functional imaging has shown that the human hippocampus is selectively activated during association of an item and the context in which it was experienced (e.g., Davachi, Mitchell, & Wagner, 2003, Ranganath et al., 2003). Thus, in rats, monkeys, and humans, a prevalent property of hippocampal firing patterns involves the representation of unique associations of stimuli and their significance, specific behaviors, and the places where these events occurred. Episodes are represented as sequences of events
Another common observation across species and many different behavioral protocols is that different hippocampal neurons fire during each successive event that composes task performance. Some cells are active during simple behaviors such as foraging for food (e.g., Muller, Kubie, & Ranck, 1987) and learned behaviors directed at relevant stimuli that have to be remembered (e.g., Hampson, Heyser, & Deadwyler, 1993), and the firing patterns have been observed across a broad range of learning protocols, from classical conditioning, discrimination learning, and non-matching or matching to sample tasks to a variety of spatial learning and memory tasks (for review, see Eichenbaum, 2004). In each of these paradigms, a substantial proportion of hippocampal neurons show time-locked activations associated with each sequential event. Many of these cells show striking specificities corresponding to particular combinations of stimuli, behaviors, and the spatial location of the event. These sequential firing patterns can be envisioned to represent a series of events, and their places that compose a meaningful episode, and the information contained in these representations, both distinguishes and links related episodes. Consider, for example, a study in which rats were trained on the classic spatial alternation task in a modified T-maze (Wood, Dudchenko, Robitsek, & Eichenbaum, 2000). Performance on this task requires that the animal distinguish left-turn and right-turn episodes and that it remember the immediately preceding episode to guide the choice on the current trial, and in that way, the task is similar in demands to those of episodic memory (Figure 9; Video 3). We found that hippocampal neurons encode each sequential behavioral event and its locus within one type of episode, with most cells firing only when the rat is performing within either the left-turn or the right-turn type of episode. This was particularly evident for cells that fired when the rat was on the "stem" of the maze, that is, when it traversed the same locations on both types of trials (Figure 9). Indeed, virtually all cells that fired when the rat was on the maze stem fired differentially on left-turn versus right-turn trials. The majority of cells showed strong selectivity, some firing almost exclusively as the rat performed one of the trial types, suggesting they were part of the representations of only one kind of episode. Other cells fired substantially on both trial types, potentially providing a link between left-turn and right-turn representations by the common places traversed on both trial types. These findings indicated that separate ensembles of neurons encoded the sequences of events that composed left-turn and right-turn trials (Figure 10). Notably, there were also some cells that fired similarly on both trial types; these might serve to link the two types of episodes. Functional imaging studies in humans have also revealed hippocampal involvement in both spatial and non-spatial sequence representation. Several studies have shown that the hippocampus is active when people recall routes between specific start points and goals, but not when subjects merely follow a set of cues through space (Hartley, Maguire, Spiers, & Burgess, 2003). In addition, the hippocampus is selectively activated when people learn sequences of pictures (Kumaran & Maguire, 2006). Even greater hippocampal activation is observed when subjects must disambiguate picture sequences that overlap, parallel to our findings on hippocampal cells that disambiguate spatial sequences (Wood et al., 2000). The hippocampus encodes events that can link related memories In virtually all the studies described above, some hippocampal neurons encode features that are common among different experiences – these representations could provide links between distinct memories. For example, in Moita and colleagues’ (2003) study of auditory fear conditioning, some cells only fired to a tone when the animal was in a particular place, whereas others fired associated with the tone wherever it was presented across trials. In the Wood et al. (1999) study on odor recognition memory, some cells showed striking associative coding of odors, whereas their match/non-match status, and places, other cells fired associated with one of those features across different trials. Some cells fired during a particular phase of the approach towards any stimulus cup, while others fired differentially as the rat sampled a particular odor, regardless of its location or match-non-match status. Yet other cells fired only when the rat sampled the odor at a particular place, regardless of the odor or its status. Still, other cells fired differentially associated with the match and nonmatch status of the odor, regardless of the odor or where it was sampled. Similarly, in Ekstrom and colleagues’ (2003) study on humans performing a virtual navigation task, whereas some hippocampal neurons fired associated with combinations of views, goals, and places, other cells fired when subjects viewed particular scenes, occupied particular locations, or had particular goals in findings passengers or locations for drop off. Also, in Rivard and colleagues’ (2004) study of rats exploring objects in open fields, some cells fired selectively associated with an object in one environment, while others fired associated with the same object across environments. The notion that these cells might reflect the linking of important features across experiences and the abstraction of common information was highlighted in recent studies on monkeys and humans. Hampson, Pons, Stanford, and Deadwyler (2004) trained monkeys on matching to sample problems then probed the nature of the representation of stimuli by recording from hippocampal cells when the animals were shown novel stimuli that shared features with the trained cues. They found many hippocampal neurons that encoded meaningful categories of stimulus features and appeared to employ these representations to recognize the same features across many situations. Kreiman, Koch, and Fried (2000) characterized hippocampal firing patterns in humans during presentations of a variety of visual stimuli. They reported a substantial number of hippocampal neurons that fired when the subject viewed specific categories of material, e.g., faces, famous people, animals, scenes, and houses, across many exemplars of each. A subsequent study showed that some hippocampal neurons are activated as a subject views any of a variety of different images of a particular person, suggesting these cells could link the recollection of many specific memories related to that person (Quiroga, Reddy, Kreiman, Koch, & Fried, 2005). This combination of findings across species provides compelling evidence for the notion that some hippocampal cells represent common features among the various episodes that could serve to link memories obtained in separate experiences. Furthermore, recent functional imaging studies have associated activation of the hippocampus in humans to the performance of transitive inference tasks similar to those described above as dependent on the hippocampus in animals. In one study, subjects learned overlapping paired associations between faces and houses or direct face-face associations (Preston, Shrager, Dudukovic, & Gabrieli, 2004). The hippocampus was selectively activated when people identified the indirect associations between faces that were paired with the same house as compared with direct face-face associations. In another study, subjects were trained on the task which involves a hierarchical series of judgments (A > B, B > C, C > D, D > E) or a series of non-overlapping judgments (K > L, M > N, O > P, Q > R; Heckers, Zalezak, Weiss, Ditman, & Titone, 2004). The hippocampus was activated when subjects performed transitive judgments as compared to novel judgments between items taken from the non-overlapping pairs. Under some circumstances, it may be possible to indirectly relate items without a memory network (O’Reilly & Rudy, 2001; Van Elzakker, O’Reilly, & Rudy, 2003), but the above described results provide compelling evidence that the hippocampus is indeed involved in binding related memories and in using these memories to make novel inferential judgments. Summing up the physiological data These various observations are consistent with the notion that hippocampal neurons in animals and humans represent the kinds of information that underlie recollection. Hippocampal neurons encode attended stimuli and behavioral actions in the context in which they occur. These representations are created for each series of events that compose a behavioral episode across a broad range of behavioral protocols and encode features of events that are shared across distinct experiences that can link memories. Spatial Functions of the Hippocampus So far this review has touched only briefly on the role of the hippocampus in spatial cognition and memory. Yet a wealth of studies have shown that the hippocampus plays an essential role in a variety of forms of spatial learning and memory in laboratory mice and rats, monkeys, and humans, as well as several other undomesticated species including fish, reptiles, bats, and a variety of avian and rodent species (e.g., Hampton & Shettleworth, 1996; Vargas, Petruso, & Bingman, 2004). Also, several studies have compared hippocampal anatomy within closely related species of rodents, birds, and bats, as well as humans with different occupations, providing fascinating evidence that hippocampal size is related to greater use of space in natural habitats (e.g., Pleskcheva et al., 2000; Safi & Dechmann, 2005; Jacobs, Gaulin, Sherry, & Hoffman, 1990; Jacobs & Spencer, 1994; Lucas, Brodin, de Kort, & Clayton, 2004; Maguire, Woollett, & Spiers, 2006). Many of these studies have been interpreted as supporting the notion that the hippocampus is selectively involved in spatial cognition and, in particular, in the creation and use of cognitive maps (Bingman, Ioale, Casini, & Bagnoli, 1988; Sherry, Jacobs, & Gaulin, 1992; Salas, Broglio, & Rodriguez, 2003; Jacobs & Schenk, 2003). However, this interpretation is challenged by several of the studies outlined above that demonstrate a critical role for the hippocampus in a variety of non-spatial memory tasks. Here I will take the view that a deeper understanding of hippocampal function in spatial cognition and memory can be had by altering the focus away from comparing spatial and non-spatial tasks, and instead focus on the fundamental demands of spatial tasks that may have led to the evolution of the hippocampus and its functions in both spatial and non-spatial memory (Sherry & Shacter, 1987). In particular, I suggest that the cognitive demands of spatial memory tasks put a heavy demand on the three fundamental features of recollection that were examined above. Many natural and laboratory spatial memory tasks involve remembering where important objects are located (e.g., food caching in birds). This demand of spatial memory is a particularly strong and common example of the general feature of recollection involving memory for items in the (spatial) context of prior experience. Other laboratory and natural spatial tasks involve remembering routes through the environment taken to find rewards or escape locations. Memories for routes provide a particularly strong example of the general feature of recollection involving memory for sequences of events, in this case events extended through space as well as time (e.g., the T-maze alternation study described above; Wood et al., 2000; for an example in humans see Shelton & Gabrieli, 2002). Yet other laboratory tasks (e.g., the Morris water maze) and many natural situations require animals to learn multiple spatial memories and interleave those memories to form a general representation of space that can be used to navigate from novel starting points (e.g., Eichenbaum, Stewart, & Morris, 1990). The networking of spatial memories and application of spatial networks (cognitive maps) in inferring novel routes (navigation) is a particularly good example of the general feature of recollection as based on linking related memories and employing the generated memory networks to make inferences from memory. From this viewpoint, many examples of spatial learning and memory in nature are especially demanding on all three of the fundamental properties of recollective memory. Thus, it should come as no surprise that the hippocampus is important for spatial memory across many species, that hippocampal neuronal activity reflects the encoding of spatial location along with other features of events (Eichenbaum, Dudchencko, Wood, Shapiro, & Tanila, 1999), and that hippocampal size relates to the high spatial memory demands for animals with larger or more complex habitats or greater demands for use of spatial memory in their everyday lives. Towards a Comparative Functional Organization of the Hippocampal Memory System
A consideration of the anatomical organization of the major circuitry involving the hippocampus and neocortex provides further insights into basic mechanisms that underlie recollection across diverse species. In primates, the hippocampus receives an enormous variety of information from virtually every cortical association area, and this information is funneled into the hippocampus via the parahippocampal region, which is subdivided into the perirhinal cortex, the parahippocampal cortex, and entorhinal cortex (Figure 11). The cortical outputs of hippocampal processing involve feedback connections from the hippocampus successively back to the entorhinal cortex, then the perirhinal and parahippocampal cortex, and finally, neocortical areas from which the inputs to the hippocampus originated (Amaral & Witter, 1995). To what extent is the organization of this system similar in mammalian species? The internal circuitry of the hippocampus itself is largely conserved across mammalian species (Manns & Eichenbaum, 2007). The subdivisions of the hippocampus are connected by a serial, unidirectional path, starting with the dentate gyrus, and continuing through CA3, then CA1, and then the subiculum. Furthermore, anatomical details involving several topographical and parallel organizations are highly similar in species including rats, cats, and monkeys, as well as other species (see Amaral & Witter, 1995 and Witter, Wouterlood, Naber, & Van Haeften, 2000 for reviews). There is also considerable conservation of the areas of the parahippocampal region. The perirhinal, parahippocampal (called postrhinal cortex in rats), and entorhinal subdivisions of the parahippocampal region are similar in cytoarchitecture in rats, mice, and monkeys, and the connectivity among these areas is also remarkably similar (Burwell, Witter, & Amaral, 1995). In contrast to the conservation of hippocampal and parahippocampal circuitry, the neocortical regions that are the ultimate origin of hippocampal inputs differ substantially from species to species. For example, there are numerous dissimilarities in the neocortex that reflect general differences between small-brained and big-brained mammals, such as cortical size, laminar stratification, and number of polymodal association areas (Krubitzer & Kaas, 2005; Manns & Eichenbaum, 2007). Furthermore, the extent of cortical areas devoted to a particular sensory modality also varies substantially between species. Despite major species differences in the neocortex, the organization of cortical inputs to the hippocampus is remarkably similar in rodents and primates. Across species, most of the neocortical input to the perirhinal cortex comes from association areas that process unimodal sensory information about qualities of objects (i.e., "what" information), whereas most of the neocortical input to the parahippocampal cortex comes from areas that process polymodal spatial ("where") information (Suzuki & Amaral, 1994; Burwell et al., 1995). There are connections between the perirhinal cortex and parahippocampal cortex, but the "what" and "where" streams of processing remain largely segregated as the perirhinal cortex projects primarily to the lateral entorhinal area whereas the parahippocampal cortex projects mainly to the medial entorhinal area. Similarly, there are some connections between the entorhinal areas, but the "what" and "where" information streams mainly converge within the hippocampus. These anatomical considerations suggest a functional organization of the flow of information into and out of the hippocampus. Here I will outline some of the functional differences between components of the parahippocampal region and subfields of the hippocampus leading to a working hypothesis about how the phenomena of recollection emerge from the organization of hippocampal pathways. Perirhinal cortex and lateral entorhinal area Substantial evidence indicates that neurons in the perirhinal cortex and lateral entorhinal cortex are involved in the representation of individual perceptual stimuli. Electrophysiological studies on monkeys and rats performing simple recognition tasks have identified three general types of responses (Brown & Xiang, 1998; Suzuki & Eichenbaum, 2000). First, many cells in these areas exhibit selective tuning to memory cues such as odors or visual stimuli. Second, some cells maintain firing in a stimulus-specific fashion during a memory delay, indicating the persistence of a stimulus representation. Third, many cells have enhanced or suppressed responses to stimuli when they re-appear in a recognition test, indicating involvement in the recognition judgment. Similarly, in humans, among all areas within the medial temporal lobe, the perirhinal area selectively shows suppressed responses to familiar stimuli (Henson, Cansino, Herron, Robb, & Rugg, 2003). Complementary studies in animals with damage to the perirhinal cortex indicate that this area may be critical to memory for individual stimuli in the delayed non-matching to sample task in rats (Mumby & Pinel, 1994; Otto & Eichenbaum, 1992) and monkeys (Suzuki, Zola-Morgan, Squire, & Amaral, 1993). These and other data have led several investigators to the view that the perirhinal cortex is specialized for identifying the memory strength of individual stimuli (e.g., Brown & Aggleton, 2001; Henson et al., 2003; Aggleton, Kyd, & Bilkey, 2004). Parahippocampal cortex and medial entorhinal area The parahippocampal cortex and medial entorhinal area may be specialized for processing spatial context. Whereas perirhinal and lateral entorhinal neurons have poor spatial coding properties, parahippocampal and medial entorhinal neurons show strong spatial coding (Burwell & Hafeman, 2003; Hargreaves, Rao, Lee, & Knierim, 2005). Furthermore, the immediate early gene fos is activated in the perirhinal cortex by novel visual cues, but fos is activated in the postrhinal cortex by a spatial re-arrangement of the cues (Wan, Aggleton, & Brown, 1999). In addition, whereas object recognition is impaired following perirhinal damage, object-location recognition is deficient following parahippocampal cortex damage in rats (Gaffan, Healey, & Eacott, 2004) and monkeys (Alvarado & Bachevalier, 2005). Similarly, perirhinal cortex damage results in greater impairment in memory for object pairings whereas parahippocampal cortex lesions results in greater impairment in memory for the context in which an object was presented (Norman & Eacott, 2005). Parallel findings from functional imaging studies in humans have dissociated object processing in perirhinal cortex from spatial processing in the parahippocampal cortex (Pihlajamaki et al., 2004). Furthermore, whereas perirhinal cortex is activated in association with the memory strength of specific stimuli (Henson et al., 2003), the parahippocampal cortex is activated during recall of spatial and non-spatial context (Ranganath et al., 2003; Bar & Aminoff, 2003). Hippocampus Compelling support for differentiation of functions associated with recollection come from within-study dissociations that reveal activation of the perirhinal cortex, selectively associated with familiarity and activity in the hippocampus as well as parahippocampal cortex, was selectively associated with recollection (Daselaar, Fleck, & Cabeza, 2006; Davachi & Wagner, 2002; Davachi et al., 2003; Ranganath et al., 2003). These and many other results summarized in a recent review suggest a functional dissociation between the perirhinal cortex, where activation changes are consistently associated with familiarity, and the hippocampus and parahippocampal cortex, where activation changes are consistently associated with recollection (Eichenbaum et al., 2006). An outstanding question in these studies is whether the parahippocampal cortex and hippocampus play different roles in recollection. In particular, the above described findings on parahippocampal activation associated with viewing of spatial scenes suggests the possibility that this area is activated during recollection because recall involves retrieval of spatial contextual information. By contrast, the hippocampus may be activated associated with the combination of item and context information. CA1 versus CA3 Several recent studies have suggested that subregions of the hippocampus may play distinct roles in memory. A particularly striking contrast comes from a comparison between two studies by Kesner and colleagues (Gilbert & Kesner, 2003; Kesner, Hunsaker, & Gilbert, 2005). In one experiment, normal rats learned associations between a particular object or odor and their locations in specific places in an open field. On each trial, one of two objects (differentiated by visual or olfactory cues) was placed at one of two locations on a large open field. If object A was in place one, a reward could be found underneath. Similarly, if object B was in place two a reward could be obtained by displacing the object. However, no reward was available if either object was presented in the alternate location. Normal animals improved in performance across days, as reflected in differentiating their latencies to approach object in rewarded vs. non-rewarded locations. Selective lesions of CA3 completely blocked acquisition of object-place associations, whereas CA1 lesions had no effect. In contrast, the opposite pattern of results was found in another study were rats were taught associations between an object and an odor that were separated by a short delay. The animals learned that if object A was presented before the delay, then a cup of sand would contain a food reward if it was scented with odor 1 (but not with odor 2). Conversely, if object B was presented first, then a cup of sand would contain a food reward if it was scented with odor 2 (but not odor 1). Memory was measured by a briefer latency to approach the scented cup on rewarded pairings (A-1 and B-2) than on non-rewarded pairings (A-2 and B-1). In normal rats, the latency to approach rewarded cups gradually decreased over daily training sessions, at about the same rate as observed in the previous object-place association study. In contrast, rats with selective CA1 lesions showed no sign of acquiring the associations between temporally separated objects, whereas rats with CA3 lesions acquired the task just as rapidly as normal animals. These results, and other similar findings (Kesner et al., 2005), are consistent with the possibility that CA3 is specialized for the representation of items in the (spatial) context in which they are experienced, whereas CA1 is specialized for representation of the order of events that are separated in time (Manns & Eichenbaum, 2005). A Species-General Model of Hippocampal Function in Recollection The diverse lines of evidence reviewed above suggest that recollection and familiarity are functionally dissociable processes, and that different components of the hippocampal system make distinct contributions to recognition memory. Evidence from neuropsychological, functional imaging, and neurophysiological studies of rats, monkeys, and humans indicate a distinct role for the hippocampus in recollection and not familiarity. The parahippocampal cortex also contributes to recollection, via the representation and retrieval of contextual (especially spatial) information, whereas the perirhinal cortex contributes to and is necessary for familiarity-based recognition. These findings are consistent with the anatomically-guided hypothesis about the functional organization of the hippocampal system presented in Figure 11 and suggest mechanisms by which the anatomical components of this system interact in support of the phenomenology of recollection (Figure 12). Following experience with a stimulus, the perirhinal and lateral entorhinal areas may match a memory cue to a stored template of the stimulus, reflected in suppressed activation that signals familiarity. Outputs from perirhinal and lateral entorhinal areas back to neocortical areas may be sufficient to generate the sense of familiarity without participation of the hippocampus. In addition, during the initial experience, information about the to-be-remembered stimulus, processed by the perirhinal and lateral entorhinal areas, and about the spatial and possibly non-spatial context of the stimulus, processed by the parahippocampal and medial entorhinal areas, converge in the hippocampus. During subsequent retrieval, presentation of the cue may drive the recovery of object-context representations in the hippocampus that, via back projections, regenerates a representation of the contextual associations in parahippocampal and medial entorhinal areas, which cascades that information back to neocortical areas that originally processed the item and contextual information. This processing pathway may constitute the subjective experience of recollection.
Conclusions This forgoing overview suggests that the phenomenology of recollection consists of the capacity to recover information about a memory that is not present at the time of testing. That information includes arbitrary associations and the context of a prior event with the cue, information about flow of events that occurred in the prior episode, and links to related memories. Notably animals as well as humans exhibit the capacity to retrieve all these forms of information derived from memory cues. Also, in animals, as in humans, the capacities to remember context, order, and related memories each depend on hippocampal function. Correspondingly, across species, neurons in the hippocampus encode the kinds of information that support these features of recollective memory. The demands for these fundamental features of recollective memory are especially evident in spatial memory performance observed across a broad variety of species. Finally, the anatomical findings and data about functional components of hippocampal circuitry are remarkably similar across mammalian species and suggest a functional organization of cortical-hippocampal pathways from which these features of recollection emerge. Because these pathways, and their particular functional roles in memory, are the same in various animal species and humans, there exists no reason to postulate a difference in the outcome of these interactions. That is, there is no reason to postulate a species difference in the capability of recollective experience, although the specific contents of memories may differ substantially because of species differences in cortical specializations. The impressive convergence of data and theory on the behavioral features of recollection and on hippocampal system organization and function offer a preliminary account of conscious recollection that involves both an evolutionary continuity and a mechanistic explanation. References Aggleton, J. P., & Brown, M. W. (2006). Interleaving brain systems for episodic and recognition memory. Trends in Cognitive Sciences, 10, 455-463. Aggleton, J. P., Kyd, R. J., & Bilkey, D. K. (2004). When is the perirhinal cortex necessary for the performance of spatial memory tasks? Neuroscience & Biobehavioral Reviews, 28, 611-24. Aggleton, J. P., Vann, S. D., Denby, C., Dix, S., Mayes, A. R., Roberts, N., et al. (2005). Sparing of the familiarity component of recognition memory in a patient with hippocampal pathology. Neuropsychologia, 43(12), 1810-1823. Alvarado, M. C., & Bachevalier, J. (2005). Comparison of the effects of damage to the perirhinal and parahippocampal cortex on transverse patterning and location memory in rhesus macaques. Journal of Neuroscience, 25, 1599-1609. Amaral, D. G., & Witter, M. P. (1995). Hippocampal formation. In G. Pacinos (Ed.), The rat nervous system, (2nd ed., pp. 443-493). San Diego, CA: Academic Press. Aristotle (350 BC). On memory and reminiscence. (J. I. Beare, Trans.). Bar, M., & Aminoff, E. (2003). Cortical analysis of visual context. Neuron, 38, 347-58. Bingman, V. P., Ioale, P., Casini, G., & Bagnoli, P. (1988). The importance of comparative studies and ecological validity for understanding hippocampal structure and cognitive function. Hippocampus, 2, 213-220. Brown, M. W., & Aggleton, J. P. (2001). Recognition memory: What are the roles of the perirhinal cortex and hippocampus? Nature Reviews Neuroscience, 2, 51-61. Brown, M. W., & Xiang, J. Z. (1998). Recognition memory: Neuronal substrates of the judgment of prior occurrence. Progress in Neurobiology, 55, 149-89. Bunsey, M., & Eichenbaum, H. (1996). Conservation of hippocampal memory function in rats and humans. Nature, 379, 255-257. Burwell, R. D., & Hafeman, D. M. (2003). Positional firing properties of postrhinal cortex neurons. Neuroscience, 119, 577-588. Burwell, R. D., Witter, M. P., & Amaral, D. G. (1995). Perirhinal and postrhinal cortices of the rat: A review of the neuroanatomical literature and comparison with findings from the monkey brain. Hippocampus, 5(5), 390-408. Clayton, N. S., Bussey, T. J., & Dickinson, A. (2003). Can animals recall the past and plan for the future? Nature Reviews Neuroscience, 4, 685-691. Clayton, N. S., Bussey, T. J., Emery, N. J., & Dickinson, A. (2003). Prometheus to Proust: The case for behavioural criteria for ‘mental time travel’. Trends in Cognitive Sciences, 7(10), 436-437. Daselaar, S. M., Fleck, M. S., & Cabeza, R. (2006). Triple dissociation in the medial temporal lobes: Recollection, familiarity, and novelty. Journal of Neurophysiology, 96, 1902-1911. Davachi, L., Mitchell, J. P., & Wagner, A. D. (2003). Multiple routes to memory: Distinct medial temporal lobe processes build item and source memories. Proceedings of the National Academy of Science, U.S.A., 100, 2157-2162. Davachi, L., & Wagner, A. G. (2002). Hippocampal contributions to episodic encoding: Insights from relational and item-based learning. Journal of Neurophysiology, 88, 982-990. Day, M., Langston, R., & Morris, R. G. M. (2003). Glutamate-receptor-mediated encoding and retrieval of paired-associate learning. Nature, 424, 205-209. Dusek, J. A., & Eichenbaum, H. (1997). The hippocampus and memory for orderly stimulus relations. Proceedings of the National Academy of Science, U.S.A., 94, 7109-7114. Eichenbaum, H. (1999). Neurobiology. The topography of memory. Nature, 402(6762), 597-599. Eichenbaum, H. (2004). Hippocampus: Cognitive processes and neural representations that underlie declarative memory. Neuron, 44, 109-120. Eichenbaum, H., & Cohen, N. J. (2001). From conditioning to conscious recollection: Memory systems of the brain. Oxford University Press. Eichenbaum, H., Dudchencko, P., Wood, E., Shapiro, M., & Tanila, H. (1999). The hippocampus, memory, and place cells: Is it spatial memory or a memory space? Neuron, 23, 209-226. Eichenbaum, H., Fortin, N. J., Ergorul, C., Wright, S. P., & Agster, K. L. (2005). Episodic recollection in animals: "If it walks like a duck and quacks like a duck…". Learning and Motivation, 36, 190-207. Eichenbaum, H., Stewart, C., & Morris, R. G. M. (1990). Hippocampal representation in place learning. Journal of Neuroscience, 10, 3531-3542. Eichenbaum, H., Yonelinas, A. R., & Ranganath, C. (2006). The medial temporal lobe and recognition memory. Annual Review of Neuroscience (in press). Ekstrom, A. D., Kahana, M. J., Caplan, J. B., Fields, T. A., Isham, E. A., Newman, E. L., et al. (2003). Cellular networks underlying human spatial navigation. Nature, 425, 184-187. Ergorul, C., & Eichenbaum, H. (2004). The hippocampus and memory for "what," "where," and "when". Learning and Memory, 11(4), 397-405. Fortin, N. J., Agster, K. L., & Eichenbaum, H. (2002). Critical role of the hippocampus in memory for sequences of events. Nature Neuroscience, 5, 458-462. Fortin, N. J., Wright, S. P., & Eichenbaum, H. (2004). Recollection-like memory retrieval in rats is dependent on the hippocampus. Nature, 431, 188-191. Gaffan, E. A., Healey, A. N., & Eacott, M. J. (2004). Objects and positions in visual scenes: Effects of perirhinal and postrhinal cortex lesions in the rat. Behavioral Neuroscience, 118, 992-1010. Gilbert, P. E., & Kesner, R. P. (2003). Localization of function within the dorsal hippocampus: The role of the CA3 subregion in paired-associate learning. Behavioral Neuroscience, 117, 1385-1394. Gothard, K. M., Skaggs, W. E., Moore, K. M., & McNaughton, B. L. (1996). Binding of hippocampal CA1 neural activity to multiple reference frames in a landmark-based navigation task. Journal of Neuroscience, 16, 823-835. Hampson, R. E., Heyser, C. J., & Deadwyler, S. A. (1993). Hippocampal cell firing correlates of delayed-match-to-sample performance in the rat. Behavioral Neuroscience, 107, 715-739. Hampson, R. E., Pons, T. P., Stanford, T. R., & Deadwyler, S. A. (2004). Categorization in the monkey hippocampus: A possible mechanism for encoding information into memory. Proceedings of the National Academy of Sciences, 101, 3184-3189. Hampton, R. R., & Schwartz, B. L. (2004). Episodic memory in nonhumans: What, and where, is when? Current Opinion in Neurobiology, 14, 192-197. Hampton, R. R., & Shettleworth, S. J. (1996). Hippocampus and memory in food storing and non-storing bird species. Behavioral Neuroscience, 110, 946-964. Hargreaves, E. L., Rao, G., Lee, I., & Knierim, J. J. (2005). Major dissociation between medial and lateral entorhinal input to dorsal hippocampus. Science, 308(5729), 1792-1794. Hartley, T., Maguire, E. A., Spiers, H. J., & Burgess, N. (2003). The well-worn route and the path less traveled: Distinct neural bases of route following and wayfinding in humans. Neuron, 37(5), 877-888. Heckers, S., Zalezak, M., Weiss, A. P., Ditman, T., & Titone, D. (2004). Hippocampal activation during transitive inference in humans. Hippocampus, 14, 153-162. Henson, R. N. A., Cansino, S., Herron, J. E., Robb, W. G., & Rugg, M. D. (2003). A familiarity signal in human anterior medial temporal cortex? Hippocampus, 13(2), 301-304. Henson, R. N. A., Rugg, M. D., Shallice, T., Josephs, O., & Dolan, R. J. (1999). Recollection and familiarity in recognition memory: An event-related functional magnetic resonance imaging study. Journal of Neuroscience, 19, 3962-3972. Hollup, S. A., Molden, S., Donnett, J. G., Moser, M. B., & Moser, E. I. (2001). Accumulation of hippocampal place fields at the goal location in an annular watermaze task. Journal of Neuroscience, 21, 1635-1644. Jacobs, L. F., Gaulin, S. J., Sherry, D. F., & Hoffman, G. E. (1990). Evolution of spatial cognition: Sex-specific patterns of spatial behavior predict hippocampal size. Proceedings of the National Academy of Sciences, U.S.A., 87(16), 6349-52. Jacobs, L. F., & Schenk, F. (2003). Unpacking the cognitive map: The parallel map theory of hippocampal function. Psychological Review, 110(2), 285-315. Jacobs, L. F., & Spencer, W. D. (1994). Natural space-use patterns and hippocampal size in kangaroo rats. Brain, Behavior and Evolution, 44(3),125-132. Kesner, R. P., Gilbert, P. E., & Barua, L. A. (2002). The role of the hippocampus in memory for the temporal order of a sequence of odors. Behavioral Neuroscience, 116(2), 286-290. Kesner, R. P., Hunsaker, M. R., & Gilbert, P. E. (2005). The role of CA1 in the acquisition of an object-trace-odor paired associate task. Behavioral Neuroscience, 119(3), 781-786. Kreiman, K., Koch, C., & Fried, I. (2000). Category specific visual responses of single neurons in the human medial temporal lobe. Nature Neuroscience, 3, 946-953. Krubitzer, L., & Kaas, J. (2005). The evolution of the neocortex in mammals: How is phenotypic diversity generated? Current Opinions in Neurobiology, 15(4), 444-453. Kumaran, D., & Maguire, E. A. (2006). The dynamics of hippocampal activation during encoding of overlapping sequences. Neuron, 49(4), 617-629. Lucas, J. R., Brodin, A., de Kort, S. R., & Clayton, N. S. (2004). Does hippocampal size correlate with the degree of caching specialization? Proceedings in Biological Sciences, 271(1556), 2423-2429. Macmillan, N. A., & Creelman, C. D. (1991). Detection theory: A user’s guide. New York: Cambridge University Press. Maguire, E. A., Woollett, K., & Spiers, H. J. (2006). London taxi drivers and bus drivers: A structural MRI and neuropsychological analysis. Hippocampus, 16(12), 1091-1101. Manns, J. R., & Eichenbaum, H. (2005). Time and treason to the trisynaptic teachings: Theoretical comment on Kesner, et al. (2005). Behavioral Neuroscience, 119(4), 1140-1143. Manns, J. R., & Eichenbaum, H. (2006). Evolution of declarative memory. Hippocampus, 16(9), 795-808. Manns, J. R., & Eichenbaum, H. (2007). Evolution of the hippocampus. In J. H. Kaas (Ed.), Evolution of nervous systems. Elsevier, Inc. (in press). Moita, M. A. P., Moisis, S., Zhou, Y., LeDoux, J. E., & Blair, H. T. (2003). Hippocampal place cells acquire location-specific responses to the conditioned stimulus during auditory fear conditioning. Neuron, 37, 485-497. Muller, R. U., Kubie, J. L., & Ranck, J. B., Jr. (1987). Spatial firing patterns of hippocampal complex spike cells in a fixed environment. Journal of Neuroscience, 7, 1935-1950. Mumby, D. G., & Pinel, P. J. (1994). Rhinal cortex lesions and object recognition in rats. Behavoral Neuroscience, 108, 11-18. Norman, G., & Eacott, M. J. (2005). Dissociable effects of lesions to the perirhinal cortex and the postrhinal cortex on memory for context and objects in rats. Behavioral Neuroscience, 119, 557-566. O’Reilly, R. C., & Rudy, J. W. (2001). Conjunctive representations in learning and memory: Principles of cortical and hippocampal function. Psychological Reviews, 108, 311-345. Otto, T., & Eichenbaum, H. (1992). Complementary roles of orbital prefrontal cortex and the perirhinal-entorhinal cortices in an odor-guided delayed non-matching to sample task. Behavioral Neuroscience, 106, 763-776. Pihlajamaki, M., Tanila, H., Kononen, M., Hanninen, A., Soininen, H., & Aronen, H. J. (2004). Visual presentation of novel objects and new spatial arrangements of objects differentially activates the medial temporal lobe areas in humans. European Journal of Neuroscience, 19, 1939-1949. Pleskacheva, M. G., Wolfer, D. P., Kupriyanova, I. F., Nikolenko, D. L., Scheffrahn, H., Dell’Omo, G., et al. (2000). Hippocampal mossy fibers and swimming navigation learning in two vole species occupying different habitats. Hippocampus, 10(1), 17-30. Preston, A., Shrager, Y., Dudukovic, N. M., & Gabrieli, J. D. E. (2004). Hippocampal contribution to the novel use of relational information in declarative memory. Hippocampus, 14, 148-152. Quiroga, R. Q., Reddy, L., Kreiman, G., Koch, C., & Fried, I. (2005). Invariant visual representation by single neurons in the human brain. Nature, 435(7045), 1102-1107. Ranganath, C., Yonelinas, A. P., Cohen, M. X., Dy, C. J., Tom, S. M., & D’Esposito, M. (2003). Dissociable correlates of recollection and familiarity within the medial temporal lobes. Neuropsychologia, 42, 2-13. Rivard, B., Li, Y., Lenck-Santini, P.-P., Poucet, B., & Muller, R. U. (2004). Representation of objects in space by two classes of hippocampal pyramidal cells. Journal of General Physiology, 124, 9-25. Roberts, W. A. (2005). Cognitive time travel in people and animals. Learning and Motivation, 36(2), 107-278. Safi, K., & Dechmann, D. K. (2005). Adaptation of brain regions to habitat complexity: A comparative analysis in bats (Chiroptera). Proceedings in the Biological Sciences, 272(1559), 179-186. Salas, C., Broglio, C., & Rodriguez, F. (2003). Evolution of forebrain and spatial cognition in vertebrates: Conservation across diversity. Brain, Behavior and Evolution, 62(2), 72-82. Shelton, A. L., & Gabrieli, J. D. E. (2002). Neural correlates of encoding space from route and survey perspectives. Journal of Neuroscience, 22, 2711-2717. Sherry, D. F., Jacobs, L. F., & Gaulin, S. J. C. (1992). Adaptive specialization of the hippocampus. Trends in Neurosciences, 15, 298-303. Sherry, D. F., & Schacter, D. L. (1987). The evolution of multiple memory systems. Psychological Review, 94, 439-469. Suzuki, W. A., & Amaral, D. G. (1994). Perirhinal and parahippocampal cortices of the macaque monkey: Cortical afferents. Journal of Comparative Neurology, 350, 497-533. Suzuki, W. A., & Eichenbaum, H. (2000). The neurophysiology of memory. Annals of the New York Academy of Sciences, 911, 175-191. Suzuki, W. A., Zola-Morgan, S., Squire, L. R., & Amaral, D. G. (1993). Lesions of the perirhinal and parahippocampal cortices in the monkey produce long-lasting memory impairment in the visual and tactual modalities. Journal of Neuroscience, 13, 2430-2451. Tulving, E. (1983). Elements of episodic memory. Oxford: Clarendon Press. Tulving, E. (2002). Episodic memory: From mind to brain. Annual Review of Psychology, 53,1-25. Uncapher, M. R., Otten, L. J., & Rugg, M. D. (2006). Episodic encoding is more than the sum of its parts: An FMRI investigation of multifeatural contextual encoding. Neuron, 52, 547-556. Van Elzakker, M., O’Reilly, R. C., & Rudy, J. W. (2003). Transitivity, flexibility, conjunctive representations, and the hippocampus. I. An empirical analysis. Hippocampus, 13, 334-340. Vargas, J. P., Petruso, E. J., & Bingman, V. P. (2004). Hippocampal formation is required for geometric navigation in pigeons. European Journal of Neuroscience, 20(7), 1937-1944. Wais, P. E., Wixted, J. T., Hopkins, R. O., & Squire, L. R. (2006). The hippocampus supports both the recollection and the familiarity components of recognition memory. Neuron, 49(3), 459-466. Waltz, J. A., Knowlton, B. J., Holyoak, K. J., Boone, K. B., Mishkin, F. S., Santoa, M. M., et al. (1999). A system for relational reasoning in human prefrontal cortex. Psychological Science, 10, 119-125. Wan, H., Aggleton, J. P., Brown, M. W. (1999). Different contributions of the hippocampus and perirhinal cortex to recognition memory. Journal of Neuroscience, 19, 1142-1148. Wirth, S., Yanike, M., Frank, L. M., Smith, A. C., Brown, E. N., & Suzuki, W. A. (2003). Single neurons in the monkey hippocampus and learning of new associations. Science, 300, 1578-1581. Witter, M. P., Wouterlood, F. G., Naber, P. A., & Van Haeften, T. (2000). Anatomical organization of the parahippocampal-hippocampal network. Annals of the New York Academy of Sciences, 911, 1-24. Wixted, J. T., & Stretch, V. (2004). In defense of the signal detection interpretation of remember/know judgments. Psychonomic Bulletin and Review, 11(4), 616-41. Wood, E., Dudchenko, P.A., & Eichenbaum, H. (1999). The global record of memory in hippocampal neuronal activity. Nature, 397, 613-616. Wood, E., Dudchenko, P. A., Robitsek, & J. R., Eichenbaum, H. (2000). Hippocampal neurons encode information about different types of memory episodes occurring in the same location. Neuron, 27, 623-633. Yonelinas, A. P. (2001). Components of episodic memory: the contribution of recollection and familiarity. Philosophical Transcripts of the Royal Society of London, Biological Sciences, 356, 1363-1374. Yonelinas, A. P. (2002). The nature of recollection and familiarity: A review of 30 years of research. Journal of Memory and Language, 46, 441-517. Yonelinas, A. P., Kroll, N. E., Quamme, J. R., Lazzara, M. M., Sauve, M. J., Widaman, K. F., et al. (2002). Effects of extensive temporal lobe damage or mild hypoxia on recollection and familiarity. Nature Neuroscience, 5(11), 1236-1241. Yonelinas, A.P., Otten, L.J., Shaw, K.N., & Rugg, M.D. (2005). Separating the brain regions involved in recollection and familiarity in recognition memory. Journal of Neuroscience, 25, 3002-3008. |
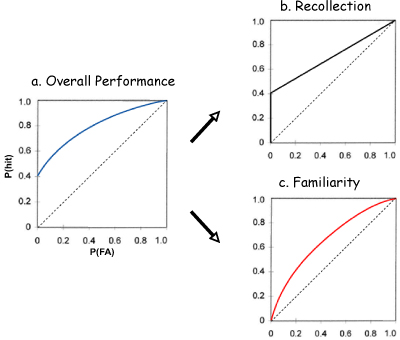
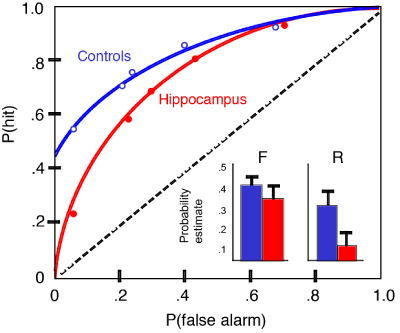
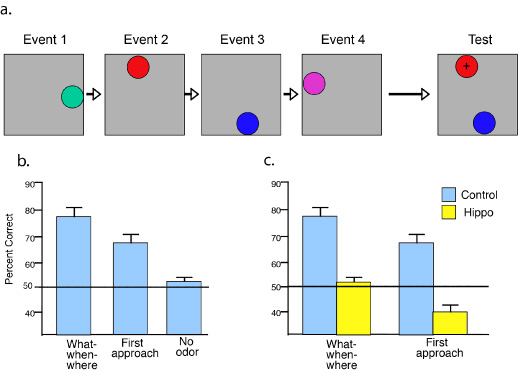
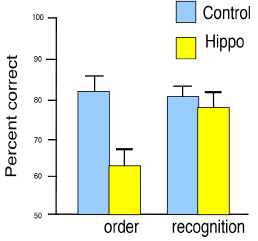
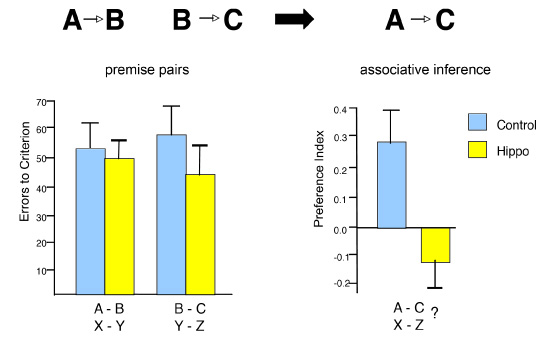
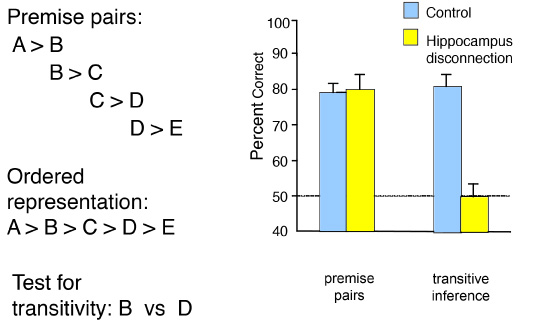

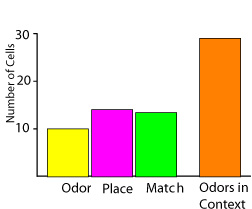
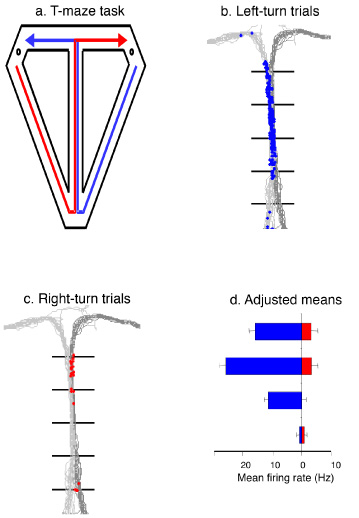
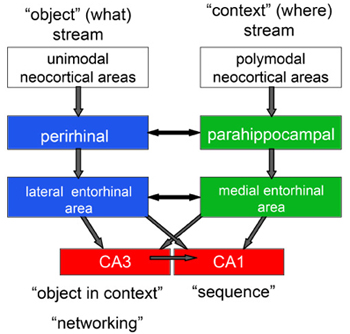
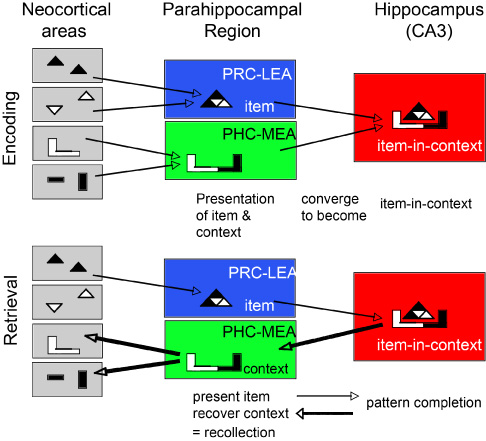
This research was supported by NIMH MH52090, MH51570, and MH71702.
|