| <<Vol. 2 Table of Contents | |
|
|
|
|
2007 |
Volume 2, pp 26-46 |

Comparative Social Cognition: From wolf and dog to humans
Enikő Kubinyi, Zsófia Virányi, and Ádám Miklósi
Eötvös University
|
|
||||||||||||||||||||||||||
|
||||||||||||||||||||||||||
|
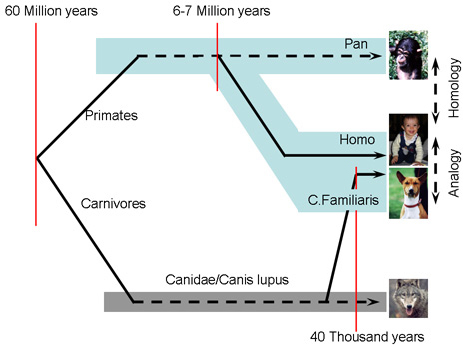
Introduction One of the most influential books on the experimental study of social behavior development was based on a 15-year-long study of dogs. Scott and Fuller (1965) argued that the study of dogs provides an understanding of developmental and genetic factors in social behavior, but additionally, because dogs evolved for living in a human dominated niche, the study of dogs has a special reference to the development of human social behavior. Accordingly dog puppies – similarly to neonates of other mammalian species – provide a good homologous model for the development of social behavior in human children. However, mainly because of the behavioral genetics approach, the experimental fell short of providing a clear ethological support for these ideas. The results had restricted relevance to those behavioral features that we today regard as parts of social cognition. The naturalistic observation of communicative and cooperative behavior between humans and dogs was lacking; humans functioned only as social reward. Finally, the dogs were maintained in conspecific family groups, mainly in kennels, and had only limited contact with humans. Thus, without denying the important contribution of these early studies to the understanding of socio-cognitive behavior in the dog, a more ethologically oriented approach is clearly needed. Dogs have three basic features, which make the species altogether unique for studying the evolution of complex social behavior. First, during evolution the behavior of dogs changed in a way that made them successful in the human social environment. Second, the behavior of dogs’ ancestor species can be reconstructed from the behavior of the wolf. Therefore, one can trace the changes that occurred during domestication that led to the emergence of a unique companion species for humans. Third, the natural socialization of dogs in the human environment offers a parallel between them and human children. Thus we suggest that the detailed ethological study of dog behavior could provide a functionally analogous model to the early evolutionary stages of human socio-cognitive behavior (Miklósi, Topál, & Csányi, 2004; Hare & Tomasello, 2005). This naturalistic model considers the dog a member of a human social group, that is, the social niche of family dogs is the human social environment. Here we report ethologically inspired behavioral observations and experimental work carried out in the mixed-species group of dogs and humans. Unique evolutionary history in the human niche Various populations of Homo sapiens sapiens left Africa in a range of 50,000-100,000 years ago (Finlayson, 2005) and moved to Southeast Asia and Europe. These were the first people to meet wolves. Based on evolutionary genetic arguments, this process may have started as early as 40,000 years ago (Savolainen, Zhang, Ling, Lundeberg, & Leitner, 2002), but archeological evidence exists only for much later events (12,000-15,000 years ago; Davis & Valla, 1978; Nobis, 1979; see Figure 1). However, this discrepancy can be explained partly by assuming that genetic - and probably behavioral - separation of the to-be-dog and wolf populations preceded the emergence of any detectable anatomical change (Vilá et al., 1997). In any case, dogs were the first domesticated animals.
Some researchers assume that domestication started with a population of wolves that became able to exploit food resources provided by humans (Coppinger & Coppinger, 2001). Later humans encouraged these wolves to join them. Subsequent selection for different behaviors and certain preferred appearances concluded with the emergence of dog breeds. It is interesting to note that dogs usually found their way to join human groups despite the variability in human social systems and cultural traditions around the world. Additionally, it is remarkable that dogs developed close contact with humans some 3,000-5,000 years earlier than any other species. As far as we know, dogs were not domesticated for any direct benefit (e.g., food). As early dog fossils from burials indicate, dogs had a special, probably partly spiritual, relationship with humans from very early on (Morey, 2006). Behavior of the ancestor Genetic research identified the wolf as the nearest evolutionary relative of the dog (Vilá et al., 1997). This fact provides a very effective comparative background, even though wolves have been distributed across the entire Holarctic and are represented by genetically and behaviorally different populations. Additionally, wolves have suffered from human hunting and environmental destruction in most parts of the world, which probably changed their behavior in many respects (e.g., increased their homophobia). Field research on wolves (e.g., Mech, 1970; Mech & Boitani, 2003) has revealed that the complexity of wolf social behavior can be compared to that observed in primates. Wolves live in closed family groups. Their dominance relationships are context-dependent (Fox, 1972); kinship relations, alliances and friendships complicate the picture (Fox, 1973). Although the general rule is that the alpha pair has an exclusive right for mating, individual preferences often override this habit (Rabb, Woolpy, & Ginsburg, 1967). Members of the pack perform cooperative behavior in hunting and when raising the offspring (Altman, 1987). Wolves possess very expressive means of communication in various visual, auditory, and chemical signals (Schenkel, 1967). All in all, the evolution of the wolf resulted in a set of complex social skills which probably contributed to the success of this species. The presence of these behavioral features provided a fortunate situation in which, by changing some aspects of their social behavior, wolves were able to adapt to the human social niche during an early phase of the modern Homo’s evolution. Given that environmental factors and experiences of the individual are comparable (see below), the differences in socio-cognitive behavior between dogs and wolves should point to those behavioral aspects that were affected during the domestication process. Naturalistic socialization with humans Socialization in the human environment can be regarded as a natural process in the dog. This does not exclude the possibility that dogs can survive and establish conspecific groups in the absence of human contact. Although we should point out that, if early human contact is provided, many individuals of even a wild species can be socialized to humans, dogs’ long evolutionary history places them in a unique position. In contrast to wolves, whose socialization to humans has to begin before day 10 (eye opening) (Klinghammer & Goodmann, 1987; Frank, Frank, Hasselbach, & Littleton, 1989), the time window for socialization in dogs (Scott & Fuller, 1965), probably closes at around 12 weeks (Freedman, King, & Elliot, 1961). Social attraction to humans develops even after 20-minute encounters twice a week, or daily eye contact with humans (Scott & Fuller, 1965). Moreover, approach tendencies toward the human cannot be diminished by punishment (electric shocks) of the puppies (Fox & Stelzner 1966) (see also below). The socialization level of dogs varies among individuals (just as is the case with children), but exposure to the human social environment is a natural consequence of their evolutionary history and not merely an experimental or procedural variable. In addition, dogs’ social environments and experiences in many respects correspond to that of children. Some data suggest that there is a close similarity between how humans interact with young infants and with dogs (Mitchell, 2001). If questioned, dog owners regard their dog (although with marked variation) not only as a member of the family (Cain, 1983) but they also felt, their relationship with their dog was similar to the relationship they maintained with their own child (Berryman, Howells, & Lloyd-Evans, 1985). Additionally, 6.5% of randomly selected veterinary center clients said they could imagine certain circumstances in which they would give a scarce drug to their pet dog in preference to a person outside the family (Cohen, 2002). Therefore, the comparison of human infants and dogs raise the possibility to investigate how two organisms with very different evolutionary paths behave after having been exposed to a similar social environment (Gomez, 2005). Family Wolf Project: Comparison of human-oriented behavior in intensively socialized wolves and dogs To date, only a few systematic comparative research programs were carried out (Frank, 1987; Zimen, 1987; Fox, 1971), but even these endeavors differed in many respects in their focus and initial working hypotheses. Further, wolf research has suffered from many methodological problems. Wolves are notoriously difficult animals to observe in the nature because their lifestyle is extremely dynamic, and they prefer remote territories (Mech, 1970). Captive animals were often forced to live under unnatural physical and social circumstances and thus provide only a limited insight into the life of this species (see Packard, 2003). Therefore, it is not surprising that there has been much disagreement on the structure and function of wolf packs and on many behavioral aspects of wolf life (for a recent review see Mech & Boitani, 2003).
From our comparative experimental perspective, further problems emerge by the various socialization procedures applied in captive wolf research. Earlier we have argued (Miklósi, Topál, & Csányi, 2004) that in order to obtain comparative experimental results, the physical and social experiences of the two species have to be at a comparable level. For this purpose two approaches seem to be obvious. As for wild wolves, we should compare them with free-ranging dogs. However, in contrast to wolves, free-ranging dog packs usually have a continuous influx from dogs that previously lived in the vicinity of humans (Boitani, Francisci, & Ciucci, 1995). Additionally, we expect that domestication affected the relationship with humans, primarily, and less affected the intraspecies social behavior. Therefore, the only reasonable possibility to reconstruct the domestication process from comparative data is to expose wolves and dogs to similar amounts of experience with humans. This approach ensures that the behavioral differences between dogs and wolves will be due to species-specific (genetic) differences, and not due to the lack of experiences (e.g., neophobia) or destructive effect of the experimental procedures. As we suggested above, humans can only socialize wolves successfully if cubs are separated from their mothers before eye opening and spend at least 20 hours in close human contact (Klinghammer & Goodman 1987; Frank et al., 1989); we decided follow this socialization regime with both with wolves and dogs. The socialization procedure Earlier studies on socialized wolves’ behavior tested problem solving, learning ability, or intraspecies aggression and did not report on human oriented behavior. In addition, the sample sizes in this work were often low (Fentress, 1967; Frank & Frank, 1982), and wolves and dogs were not socialized to comparable levels (Hare, Brown, Williamson, & Tomasello, 2002). We decided to avoid these insufficiencies. In the years 2001-2003 we obtained 13, 4 to 6 day-old wolf cubs from 5 litters (6 males and 7 females in all) and 11 dog puppies of similar ages from 5 litters (6 males and 5 females; all mongrels, born in shelters). The unique feature of this program was that each cub and puppy had its own human caretaker, who spent 24 hours a day together with the animal for a period of about 9-16 weeks (Figure 2). Although the animals had the chance to meet conspecifics regularly (at least weekly), they spent most of their time in close contact with the human caretaker. Additionally, every caretaker took special care to avoid competitive situations and aggressive interactions in order to maintain a friendly and trustful relationship with the animal. The caretakers often carried the animals on their body in a pouch, and they slept together at night. The animals were fed first by bottle, then later (from the age of 4-5 weeks) with solid food by hand. When the subjects’ motor activity made it possible, they were trained to walk on leash and execute some basic obedience tasks. The caretakers carried the cubs and the puppies to various places either by car or public transport. Also, they were regular visitors at the university, participated in dog-camps, and frequented dog training schools (Figure 3). From their 3rd week of life the animals were tested regularly (from 3 to 9 weeks, weekly, except at the 7th week) in a laboratory at the university. These behavioral experiments tested for social preferences, social and physical neophobia, reaction to dominance, retrieval of objects, communication with humans, and possessivity. After this intensive period, the wolves were gradually integrated into a wolf pack at Gödöll (near Budapest), and the caretakers visited them once or twice a week. These visits included social contact and joint exercises (going for a walk), training of obedience tasks (sitting and lying down for food reward), and experimental work, which was terminated when the wolves became 1.5-2 years old. The dogs received the same intensive socialization procedure as the wolves, but they either stayed with their caretaker, or we found adoptive caretakers for them.
Tractability of our intensively socialized wolves Due to the intensive socialization, subsequent training, and testing, it was easy for the caretakers to handle the wolves even after the juvenile period. To demonstrate this, we examined the behavior of 8 wolves (1 male and 2 females, aged 2 years; 4 females and 1 male, aged 1 year) in a set of tractability tasks (Ujfalussy, Virányi, & Kubinyi, 2003). The other 5 intensively socialized animals were not available at the time of testing; however, they were kept and treated in the same way as their mates. The animals were observed only once in the test battery presented below, therefore the data are only for giving an indication. Response to calling by name in the pack. The animals were kept together as a pack, with other (not intensively socialized, 3- or 4-year-old) individuals in a 10,000 m2 enclosure. The caretaker stepped in the enclosure, shouted her animal’s name several times, and tried to touch the approaching wolf. All animals responded to the call and approached the caretaker in less than 46 seconds (latency of the first touch from the first call: mean (sec) ± SE = 22.3 ± 5.3). The caretaker held the wolf, put a collar and leash on it, and the caretaker and wolf left the enclosure. During the subsequent tasks, the tested animal was separated from its mates in a kennel, where the caretaker and a cameraman that the animal was familiar with were present. Sitting and lying down. The caretaker instructed the wolf to sit and to lie down twice. The caretaker had a food pellet in her hand, held her arm above the wolf, and said "Sit!" in Hungarian. When the wolf sat down, the caretaker crouched, and said "Lie down!" in Hungarian. Every wolf performed the tasks. Then a woman (aged 23) with whom the wolves were not familiar repeated the same actions. Every wolf without exception performed the tasks again. The sitting down task was repeated by both persons. Now the aim was to keep the wolf sitting as long as possible. The length of time spent sitting during the 2 trials was averaged (caretaker: mean (sec) ± SE = 10.9 ± 2.2; unfamiliar woman: mean (sec) ± SE = 8.7 ± 2.1). There was no difference between obeying the two persons (t 5 = 2.1, p = 0.09).Wearing dog accessories. The wolves were accustomed to wearing muzzles; therefore, it was not surprising that every animal let the caretaker put a muzzle on them, without any sign of disagreement, in less than 15 seconds (mean (sec) ± SE = 10.5 ± 1.6). Similarly, all of the wolves allowed the caretaker to put dog socks on one of their front and one of their hind paws, although they had never had such an experience before (mean (sec) ± SE = 101.8 ± 34.3). Only 2 animals pulled off the socks in the subsequent 30 seconds. However, when the unfamiliar woman tried to put a collar on the wolves, 4 animals (from both age-groups) showed aggressive signs, so we stopped the test. The other 4 animals accepted the well-known collar from the person they were not familiar with (mean (sec) ± SE = 33.5 ± 6.8). Social and physical neophobia. The unfamiliar woman invited the wolf to play with a toy. Each animal approached her, 2 played with the toy, 2 were neutral, and 4 (all females) showed the sign of mild stress (held their tail between their legs). In another neophobia test, we observed how our animals responded to a 1 m high flag barrier (10 cm wide red strips are attached to a string with a 10 cm separation between them). This kind of barrier was commonly used to hunt or live-trap animals in Eastern Europe. The general experience is that unsocialized wolves are afraid of passing through such a barrier (Musiani et al., 2003). However, 6 out of our 7 wolves could be enticed to pass through this flag barrier within a minute. A 2-year-old female crossed it without any kind of encouragement. Overall, our wolves could be handled similarly to dogs in many ways, with moderate individual differences (Ujfalussy et al., 2003). Therefore, intensive early handling proved to be an effective means of socializing wolves to a level comparable to dogs, which made comparative behavioral experiments possible. Previous attempts at intensive socialization of wolves were usually restricted to a single animal (e.g., Fentress, 1967), which made comparative work questionable. Additionally, some comparative work involved either less socialized wolves than the dogs that were used for comparisons (e.g., Hare, Brown, Williamson, & Tomasello, 2002) or deliberately utilized a mixed socialization procedure. For example, the wolves reared by Frank & Frank (1982) spent 12 hours with their mother and littermates and 12 hours with the experimenters and 1 littermate between the 11th day and 6th week. After following this protocol, Frank et al. (1989) reported that their wolf cubs were nearly as wary of humans as were cubs reared without human contact. Yet, the most frequently used protocol is to take away every cub from the mother before the 10th day, keep the cubs together, and bottle-feed them in the first few weeks. After two months the wolves usually live in a kennel as a pack (Feddersen-Pettersen, 1991; Hare, Brown, Williamson, & Tomasello, 2002). In contrast, we studied a fairly large sample of wolves that were separated both from their mother and their littermates on days 4-6 and stayed with humans for up to 3-4 months. For comparative purposes, we also studied a group of intensively socialized dogs. In our view, this represents an important first step in comparing the social behavior toward humans of wolves and dogs. Dog Behavior From a Comparative Perspective Early comparative projects had various research agendas, which often led to paradoxical results. For example, when the famous German zoologist, Bernard Grzimek (1942), proposed that domestication might have enhanced the dog’s cognitive abilities, the reason behind this was probably – at least partly – due to his bad experiences in training socialized wolves. In subsequent experiments, he found that his dogs performed better at recalling the location of cached food than wolves did. In contrast, Hemmer (1990) assumed that the domestication process had a detrimental effect on cognitive abilities of dogs (and other species) because the environment provided by humans buffered these species against selective forces that have an effect on natural populations. Frank (1980) hypothesized that the main difference between wolf and dog lies in the way they process information. According to him, wolves possess an instinctual system, which is responsible for behavioral traits needed for survival, while the cognitive system controls complex behavioral skills needed for social life and hunting. Evolution merged these two systems in dogs, which paved the way for a more flexible behavioral organization that reacts to a much wider range of stimuli and shows a more flexible behavioral adjustment in comparison to their ancestor. We would not deny that there are many interesting ideas in these approaches, but given the very limited comparative experimental evidence, a definite theory on the underlying behavioral and cognitive machinery is premature. As a working hypothesis, we assume that dogs’ socio-cognitive abilities were shaped by human social setting (see above); therefore, we categorize the behavior elements under investigation in accordance with the scheme of Human Behavior Complex (HBC) developed by Csányi (2000). HBC has three main dimensions: sociality, behavioral synchronization, and constructivity. One advantage of this approach is that it decomposes complex social behavior into smaller traits which can be investigated separately in a comparative perspective. The value of the comparative model will depend on to what degree the counterparts of the HBC complex can be experimentally tested and evaluated in different species. Based on the HBC we have introduced the Dog Behavior Complex (DBC; Topál, Miklósi, et al., 2006). Here we cover those elements of DBC that have a particular relevance from a wolf-dog comparison perspective.
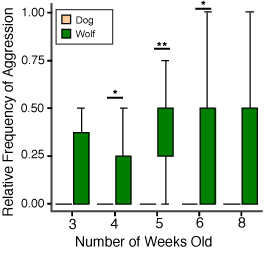
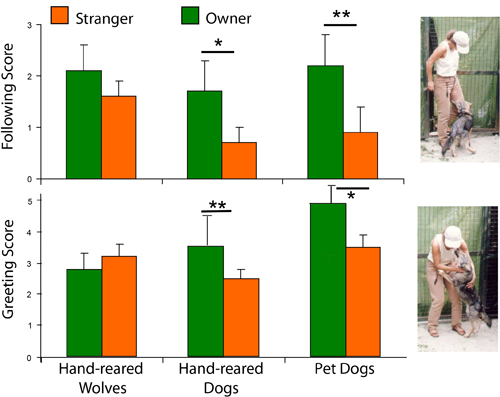
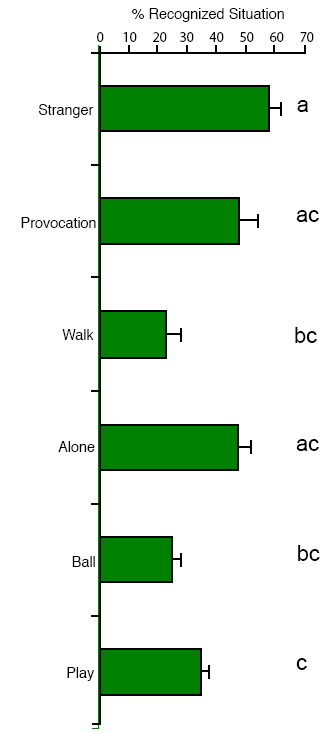
Socialization and social attraction Early social preference. Wolf cubs socialized by humans show species specific affective behaviors toward their caregivers, but whether this reflects a change in preference (relative to conspecifics) was not tested. Frank and Frank (1982) found that wolves exposed both to conspecifics and humans showed a preference for the canid partner if they were offered a choice. This was in contrast to Malamute pups, which displayed more signs of preference for humans, and although they maintained their relationship with their mother, the dogs rapidly developed strong emotional ties with humans. However, recently, we have shown that intensively socialized wolf cubs did not prefer to stay in the proximity of a conspecific at the age of 3-5 weeks if their caretaker was also present (Gácsi et al., 2005; Figure 4). This result indicates that more intensive socialization than that of Frank and Frank (1982) can inhibit preference toward conspecifics, at least at this early age. By way of comparison, hand-reared dogs were more vocal, approached humans more frequently, wagged their tails, and looked at the human’s face, which was not observed in wolves (Gácsi et al., 2005). These interesting results do not answer all questions about dogs’ social preferences. We know from previous experiments that dogs easily develop preferences for other species, such as rabbits (Cairns & Werboff, 1967) and cats (Fox, 1970), if raised together with these species early in life. Therefore, it is not sure whether dogs’ preference for humans is the result of a special selection in the human environment, or whether dogs have a decreased specificity for accepting heterospecifics as group mates. Also, dogs and wolves are not the only species that can be disposed toward accepting humans as social partners. Selection for tameness affects the socialization periods and the predisposition towards humans in silver foxes (Belyaev, 1978; Plyusnina, Oskina, & Trut, 1991). Aggressive behavior. Wolves are often characterized as aggressive animals, based upon the behavior in captive packs, but experienced field observers describe them usually as relatively peaceful, and increased aggression is restricted mainly to certain periods of social life (e.g., mating season; Mech, 1999; Packard, 2003). Given the wide range of aggressive behavior in dogs, the hypothesized overall decrement of aggressive behavior in dogs is difficult to support experimentally. Interestingly, long-term observation of wolves and dogs (Poodles) during their first year of life proved that juvenile wolves were less aggressive (Feddersen-Pettersen, 1991). Similarly, Frank and Frank (1982) comparing wolves and Malamutes could not find enhanced aggressiveness in cubs of the former species. We conducted several behavioral experiments weekly in a laboratory between the first 3-9 weeks of the animals’ lives. Tests usually began when the animal was placed on a starting point by a familiar experimenter. After the test the experimenter picked up the subject and handed it to the caregiver. None of the dog puppies behaved aggressively in the 131 interactions with the familiar experimenter during the tests, but among the 13 wolves there were 9 individuals that growled at and/or tried to bite the experimenter (in 41 cases out of 143 interactions or 29%). The greater number of growls and attacks (e.g., attempted biting) in wolves and the absence of these behaviors in the dogs could be best explained by supposing that wolves either did not like to be touched or constrained in their movements, or they had a lower threshold for the elicitation of aggressive behavior (Gyri, 2004; see also Gácsi et al., 2005; Figure 5). We cannot report on aggressive behavior of our wolves at an adult age (except at the Tractability tests, see above) because we stopped the experimental work when the wolves reached the age of 2. Before that age wolves did not attack either their caretaker or other familiar experimenters, although they displayed agonistic behavior several times towards them. Knowing well that hand-reared wolves can pose a real threat to humans (e.g., Fox, 1971, pp. 102-109) handlers tried to preclude conflicts if it was possible. Attachment to the human caregiver. Earlier work has established that dogs show attachment behavior to their caregivers analogous to that of human toddlers in the Strange Situation Test (Topál, Miklósi, Dóka, & Csányi, 1998). The original test (Ainsworth & Wittig, 1969) consisted of several episodes of separation and reunions between a mother and child. Based on this work, the functional definition of attachment is that the subject is able to discriminate and respond differentially to the object of attachment, shows specific preference toward the caregiver (proximity and contact seeking), and responds to separation from and reunion with the attachment figure differently than to similar experiences with others (see Gácsi, Topál, Miklósi, Dóka, & Csányi, 2001). Dogs, like children, show preference for their caregivers and emit specific greeting behavior in comparison between the caregiver and a stranger (Topál et al., 1998; Gácsi et al., 2001; Marston, Bennett, & Coleman, 2005). Comparing the attachment behavior of our 4-month-old hand-reared wolves and dogs revealed marked differences. Puppies showed an adult-like pattern of attachment behavior that can be characterized by more proximity and contact seeking toward their caregiver in comparison to an unfamiliar person, whereas such discrimination was lacking in wolf cubs (Topál, Gácsi, et al., 2005; Figure 6). This occurred despite the fact that the social experience with humans was comparable for both species. If attachment behavior is the outcome of social experience only, we would not expect a difference between dogs and wolves. Thus the lack of attachment toward the primary caregiver (who bottle-fed them and with whom they spent their first week in close contact) in wolves is likely to have genetic basis. And by similar logic, the attachment behavior of dogs is probably the result of their evolutionary adaptation to the human niche.
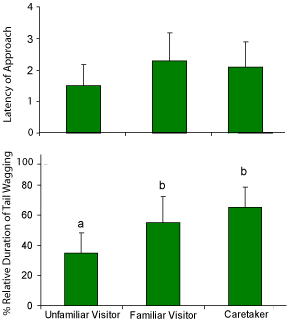
The lack of attachment in wolves does not mean that wolves do not recognize and prefer their primary caregiver in comparison to other familiar humans or an unfamiliar person. In a free interaction situation, after longer separation from the caregiver (several days), we observed the greeting behavior of the wolves. When different persons entered the enclosure (one by one) the wolves approached their caregivers sooner than others, wagged their tail more often in their presence (note that at the age of 3-5 weeks we could not register tail wagging on the same subjects in the object preference test, in contrast to dogs). Also, wolves jumped up on their caregivers more frequently compared to others. Interesting to mention here, and telling about the memory capacities of wolves, they approached an unfamiliar person sooner at their first meeting than at their second, although sometimes several weeks passed between the two occasions (Virányi et al., 2002; Figure 7).
Cooperation. Wolves are famous for their cooperative ability when hunting large game in packs (Mech, 1970). They employ a range of tactics to chase, catch, and kill their prey (Peters, 1978), although experimental modeling of these complex abilities is not yet available. Interestingly, data are also scarce in the case of the dog. It seems that the domestication process disrupted the ancient social organisation. In feral dogs, paternal and alloparental care are reduced; they do not hunt cooperatively, but prefer scavenging or hunting small prey alone (Boitani et al., 1995; Butler, du Toit, & Bingham, 2004). However, it is obvious that a gun dog or an assistant dog shows cooperative behaviour in the context of the human-dog relationship. In a descriptive study we found complex division of labor when we observed the interaction between blind persons and their trained guide dogs (Naderi, Miklósi, Dóka, & Csányi, 2001). When they navigate together, certain tasks can be executed by one partner only. The human knows the goal of the trip, and he/she might even go along a special route. In contrast, only the dog receives visual information about the actual state of the environment. A detailed behavioral analysis revealed that human and dog alternatingly took on the role of initiation during walking. For most of the time one partner led the other in one or two subsequent actions and then the initialization was handed over to the other partner. Such a finely tuned cooperative interaction between dogs and humans, where the initialization of actions is exchanged continuously, can be regarded as a functional equivalent of complex forms of between-human cooperation characterized by Reynolds (1993) as complementing. Synchronizing function of behavior: Learning by observation Successful functioning of social groups often depends on the ability of individuals to carry out actions in a coordinated way. The human species is especially inventive from this point of view because of our inclination for music and dance, and the ability of empathy or hypnotizability are regarded to serve this function (Csányi, 2000). However, there are several behavioral mechanisms that support the group-level synchrony in animals, for example, learning by observation. Davey (1981) suggests that learning by observation facilitates social cohesiveness. Social learning provides the advantage that behavioral skills can be acquired with relatively little effort by observing a knowledgeable individual (demonstrator). Synchronization is achieved at the group level because social learning facilitates the distribution of behavioral skills among the group members and between generations. In spite of the fact that in the case of many social mammals social learning has been shown to exist, very little is known about the Canids. Although, in reviewing the situation from an ecological perspective, Nel (1999) argued for the role of social learning in these species, experimental evidence was not presented. Frank (1980) cited some anecdotal evidence that wolves could find out how to open latches of their cage by observing humans, in contrast to the dogs in adjacent kennels, which could not do this. In his view, this suggested that wolves are superior to dogs with regard to social learning. While there is still limited evidence of social learning in wolves, recent experiments provided clear support for such ability in dogs, which refute Frank et al.’s (1989) suggestions that dogs are not able to learn by observation. Dogs that are exposed to a human or conspecific demonstrator solving detour or instrumental tasks are more successful at those tasks than dogs that have not witnessed the demonstration (Pongrácz et al., 2001; Pongrácz, Miklósi, Tímár-Geng, & Csányi, 2003; Pongrácz, Miklósi, Timár-Geng, & Csányi, 2004). Moreover, dogs can use socially obtained information for overcoming their own habitual action (Pongrácz, Miklósi, Kubinyi, Topál, & Csányi, 2003), or adopt a virtually useless habit (Kubinyi, Miklósi, Topál, & Csányi, 2003) by following some pattern of human demonstrations, even in the absence of any direct reward (Kubinyi, Topál, Miklósi, & Csányi, 2003). More recently, some evidence has been obtained that dogs could be capable of using a human behavior action as a cue for showing a functionally similar behavior (Topál, Byrne, Miklósi, & Csányi, 2006) and follow social rules in the context of interacting with humans (Topál, Kubinyi, Gácsi, & Miklósi, 2005). In sum, dogs proved to be very flexible in social learning in that they are able to learn from (members of) another species and do so without food or any other causal reinforcement, even in cases where the goal or the result of the action is not clear. This ability also emerges in human development (e.g., Meltzoff, 1996). It is unfortunate that such experiments have not been carried out on socialized wolves. Constructivity: Communicative behavior Understanding of human gestural signals. Many assume that the increased communicative abilities of dogs contributed to their successful integration into the human niche. However, little attention was directed to the heterospecific aspects, as experimental research was devoted mainly to intraspecific communication (wolves: Fox, 1971; dogs: Bradshaw & Nott, 1995). Some researchers have proposed that the ability of dogs to communicate with humans is mainly based on the complex communicative skills of wolves. For example, wolves often communicate with each other by pointing to a phenomenon of interest, when, for example, they indicate the direction of prey for the others (this behavior was probably selected for in pointers). The use of such a directional cue could, in principle, facilitate the understanding of human directional signals, such as pointing. Recently, different experiments have been conducted to compare the ability of socialized wolves and dogs to find hidden food on the basis of human signals. Hare, Brown, Williamson, and Tomasello (2002) tested 7 family dogs and 7 sanctuary-kept wolves’ reaction to proximal pointing and gazing cues while standing in front of their cage. In contrast to dogs, wolves did not rely on human gestures. However, we should note that although the wolves were hand-reared, the littermates had not been separated, and after the first two months they spent their life in an enclosure. Their experiences were hardly comparable to those of family dogs (see also Packard, 2003). The rearing history influences the performance: Human-reared wolves outperformed mixed socialized wolves (reared both by humans and a wolf) in a visual discrimination task (Frank et al., 1989). It is not surprising that our wolves, which were socialized to humans at a comparable level to dogs, were able to find food spontaneously on the basis of simple discriminative human cues similar to those mentioned above. Wolves performed above chance if the experimenter tapped the bowl, presented proximal pointing (when the pointing finger is at 10 cm from the bowl), or stood behind the bowl containing the food, even in the lack of gazing at the baited bowl in all cases (Miklósi et al., 2003; Virányi et al., 2006a; Figure 8). These results suggest that in Hare et al.’s study (2002) the low performance of wolves could be due to their insufficient level of socialization.
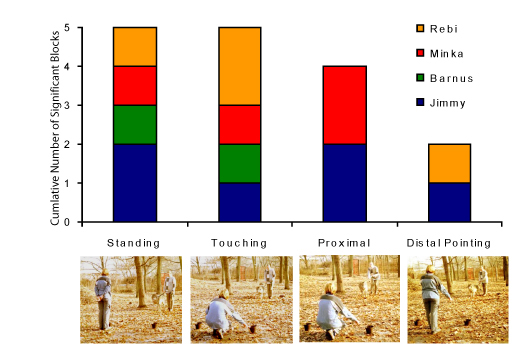
Another important, but often neglected, aspect of pointing tasks is that when the experimenter is pointing, the animal needs to look at the human (body and face) instead of looking elsewhere. Interestingly, even our extensively socialized wolves did not search for eye contact with humans spontaneously either in this or in other experimental situations (Virányi et al., 2006a; Miklósi et al., 2003). This would partially explain why wolves performed relatively poorly with more subtle gestures, such as momentary distal pointing. (In momentary distal pointing, the experimenter’s pointing finger is more than 50 cm from the bowl, and he/she moves his/her arm to the initial position after pointing, so the subject cannot see the cue when approaching the bowl. The experimenter looks at the animal and not at the bowl.) Importantly, dog puppies as young as 4 months old are able to perform well with the momentary distal pointing, without any special, intensive, and early socialization to humans. Although the everyday life of dogs provides many opportunities to learn about human gestures, dogs proved to rely on the distal pointing cue at the same level at the ages of 4 and 11 months. After extensive training, wolves significantly improved in parallel with increased readiness to look at the pointing human. At the age of 11 months, wolves reached the performance level of same aged, naïve dogs (Virányi et al., 2006a; Figure 9). It is important that wolves showed large inter-individual variations in their performance. This high variability could be the base of selection for improved communicative abilities toward humans that dogs seem to be strongly selected for. In human evolution, communicative behaviour had a special importance (Donald, 1991; Csányi, 2000); thus it is probable that the domestication process affected dogs’ ability to communicate with humans. In fact, although there is still debate about whether chimpanzees understand the pointing gesture (Miklósi & Soproni, 2006), it is now known that dogs can reach similar levels of success at pointing as 1.5-2-year-old children (Lakatos, Soproni, Dóka, & Miklósi, submitted manuscript). Dogs are able to respond to highly various forms of pointing gestures, such as pointing with one’s leg (Soproni, Miklósi, Topál, & Csányi, 2002) and are able to recognize minute behavioral cues characterizing human visual attention (Call, Bräuer, Kaminski, & Tomasello, 2003; Gácsi, Miklósi, Varga, Topál, & Csányi, 2004; Virányi, Topál, Gácsi, Miklósi, & Csányi, 2004).
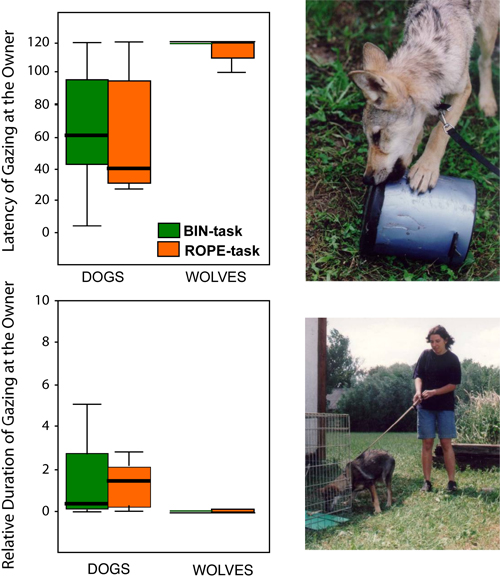
Initiation of communicative interactions. Discussing the understanding of human gestures, we noted that wolves had decreased willingness to look at humans even after intensive socialization. This is not the case in dogs, which look at humans in cases of uncertainty (Topál, Miklósi, & Csányi, 1997) and when they are physically constrained from obtaining hidden food (Miklósi, Polgárdi, Topál, & Csányi, 2000). Wolves in this latter situation alternated their gaze significantly less between the location of food and their caretaker than dogs did (Virányi et al., 2006b). The result that wolves are not ready to initiate communicative interactions with humans gained more support from other comparative experiments. We designed a test situation consisting of two phases. In the first phase the subjects learned how to obtain a piece of meat by pulling a rope. Importantly, there was no difference either in the speed of learning or in the orientation of scratching and sniffing the apparatus between the two species (in contrast to Frank & Frank, 1982). For the second phase the animals were prevented from obtaining the reward because the rope was fastened imperceptibly to the frame of the apparatus. We wanted to know whether there is any difference in the behavior of dogs and wolves in such an insolvable situation. The analysis found that after some unsuccessful attempts the dogs looked at the human caretaker standing near to them, but the wolves ignored the human, kept on trying on their own, or gave up and had a rest. The results were the same when a food item was in a covered litter bin, and after some successful trials, we attached the lid to the container in such a way that it could not be opened (Miklósi et al., 2003; Figure 10). Frank and Frank (1985, p. 271) also noted the difference between the two species in following gazing signals: "The wolves generally attacked each puzzle immediately upon release from the start box and persisted until either the problem was solved or time had run out. In contrast, the malamutes investigated puzzle boxes only until they discovered that the food was not easily accessible, after which they typically returned to the start box and performed a variety of solicitation and begging gestures toward Experimenter 1."
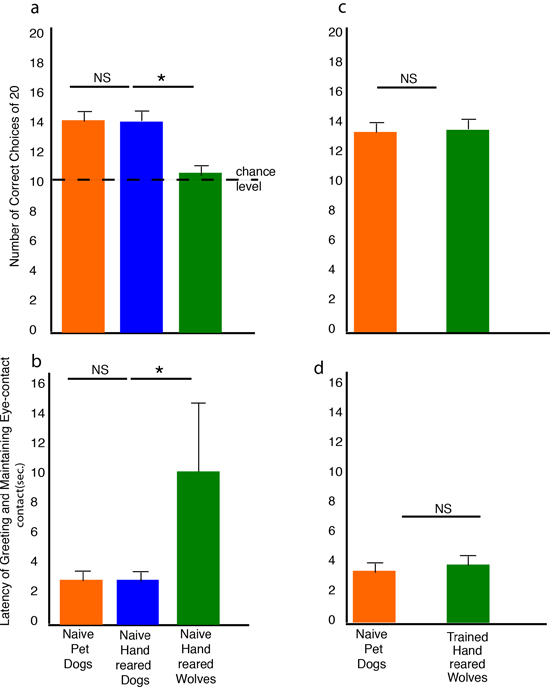
In a preliminary study we observed readiness for eye contact by means of operant conditioning. Subjects at the age of 5 and 9 weeks were rewarded immediately with a food pellet if they established eye contact with a familiar experimenter. Five-week-old wolves, although motivated by food, fell asleep during the 4-min test-session: a strategy they employed for dealing with unsolvable problems in other tests as well. Four weeks later, the wolves made intensive attempts to reach the plate containing the food as the session went on, but the frequency of their gazing at the experimenter did not change. In contrast, dogs established eye contact with the experimenter significantly more frequently during the fourth minute (Gácsi et al., 2005). Subsequent tests on our intensively socialized wolves and dogs confirmed that dogs appear to have an innate readiness to look at human faces, whereas wolves seem to ignore the human gaze. We observed the behavior of 9-week-old wolf and dog puppies called to a closed coop containing fresh chicken meat. The caretaker, standing at 3 m from the coop, called the animal by name. Dogs looked at their caretaker sooner than wolves. Both species’ motivation for obtaining the food was probably similar, since both dogs and wolves orientated to the coop for about same amount of time. We repeated the test at the age of 6 months, but now the caretakers were not allowed to talk to the animals and stood motionless 2 m from the coop for 2 minutes. Again, dogs looked at their caretakers sooner and for longer than wolves. However, wolves orientated more to the coop than dogs did. At the time of testing wolves already lived together in a pack for more than 2 months, which probably had an effect on their competitiveness, motivation, and endurance (Újváry, 2004). Two nonexclusive processes might account for the difference between dogs’ and wolves’ willingness to look at a human face. Dogs may have been selected for an inclination to look at humans. Also, or alternatively, dogs may tolerate being gazed at by humans better than wolves. Extended gazing by humans may act as a threat to wolves (see Vas, Topál, Gácsi, Miklósi, & Csányi, 2005) which induced subordinate or agonistic behavior (Fox, 1971; Schenkel, 1967). In human communication, the dynamics of eye (or face) contact is understood as initialization and maintenance of a communicative interaction (Gomez, 1996). We assume that humans selected for this human-like communicative behavior in the ancestors of modern dogs. This relatively subtle change in behavior could have crucial consequences, as it provides a potential starting point for the integration of dog and human communication systems (Miklósi et al., 2003, p. 765). In other words, looking at our face, a dog can modify our behavior in a way that is advantageous for the animal. This behavior can be interpreted as social tool use (Gomez, 1996).
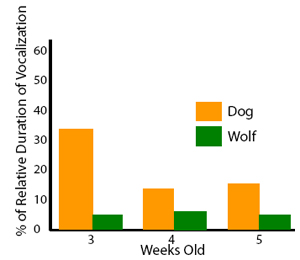
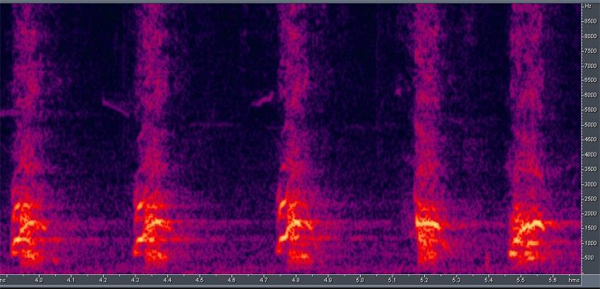
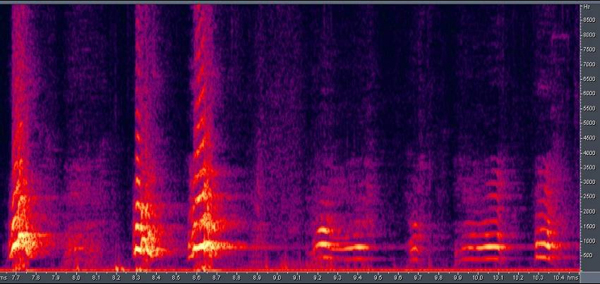
Vocal communication. Barking is one of the most striking features of dog behavior, yet it has received little attention. Ethological observations has provided evidence that barking is also part of the wolf’s vocal repertoire (e.g., Schassburger, 1993), but it is used only with a very restricted meaning of warning or protesting with relatively little variation in acoustic structure (Feddersen-Pettersen, 2000). In contrast, dogs bark in a variety of different situations and produce a highly variable vocalization (Fox, 1971; Feddersen-Pettersen, 2000; Yin, 2002). Quantitative differences in vocalizations start at an early age. Observing 3-5-week-old human-fostered wolf cubs and dog puppies in a series of object-preference tests revealed that puppies spent more time vocalizing during the tests than cubs did. However, we never observed growling or barking. All vocalizations were high pitched sounds (e.g., whining or yelping), mainly when the animals were not in the proximity of the objects. We infer that the behavior is a sign of distress (Gácsi et al., 2005; Figure 11). Earlier theories of dog vocalization suggested that it simply represents a hypertrophied behavior which was a by-product of domestication, but recent studies have shown that the situation might be more complex. Human subjects exposed to bark sequences recorded in different behavioral contexts (e.g., a dog is tethered to a tree, attacks a stranger through a fence, or plays with its caretaker) were able to categorize these contexts correctly (significantly above chance) when provided with a list of possibilities (Pongrácz, Molnár, Miklósi, & Csányi, 2005; Figure 12 and 13). In a subsequent test, subjects indicated the emotional content of the bark using a list of adjectives. For example, barks recorded when the dog attacked the stranger received the highest scores on aggressiveness. The analysis of the barks revealed that various acoustic parameters correlated with the assumed emotional content. Barks recorded in aggressive situations were characterized by lower fundamental frequency and reduced harmonic content with a shorter inter-bark interval. Affiliative vocalizations had a higher fundamental frequency and included more harmonics (Pongrácz et al., 2005; Figure 12). This suggests that selective processes during their socialization among humans appears to have favored dogs able to signal their situation and their emotional states. This change in the use of barking seems to correspond to the structural rules suggested by Morton (1977). These hypothesized changes in barking together with similar changes in the visual communicative system of dogs suggest a trend toward an increase in the number and kinds of communicative behaviors in dogs. Such increased flexibility in communicative behavior, which was also noted by Frank (1980), would be highly adaptive in forming complex communicative interactions with humans.
Overview on the Wolf-Dog Comparative Data
Raising wolf cubs and dog puppies in an identical way revealed many specific social behavioral differences between the two species, especially with regard to their interactions with humans. Even at an early age (3-5 weeks), dogs displayed more communicative signals (e.g., vocalization, tail wagging, gazing at the human’s face) and were less aggressive and avoidant than wolves, although the general activity level did not differ between the two species (Gácsi et al., 2005). Due to human fostering, 5-week-old wolves showed a clear preference for their caregiver in an object preference test, if the other stimulus-object was another human (Gácsi et al., 2005; Figure 4). However, in contrast to dogs, wolves’ preferences for the caregiver did not develop into a behavioral pattern that could be categorized as attachment. Hand-reared dogs and pet dogs, but not individually socialized, hand-reared wolves, exhibited highly different responsiveness to their caregiver compared to an unfamiliar human as early as at the age of 16 weeks (Topál, Gácsi, et al., 2005). While wolves did not display characteristic patterns of attachment toward their caregiver, their preference for her remained strong at the age of 1 or 2 years (Virányi et al., 2002). Many assume that domestication affected dogs’ ability to communicate with humans. Wolves, given that their socialization is comparable to that of dogs, were able to follow human cues that have a local enhancement or food-hand association component (e.g., touching, proximal pointing; Miklósi et al., 2003; Virányi et al., 2006a). Recent results for farm-reared foxes not selected or trained at approaching humans (Hare et al., 2005), show that they too were able to follow human proximal pointing and gazing (Hare et al., 2005). In this context it is important to recall that our hand–reared dogs, but not wolves, were able to use more difficult human pointing gestures (e.g., momentary distal pointing) spontaneously and that wolves needed more training to reach the same level of success that dogs reached instantly. The reason for this difference might be that in contrast to dogs it was very difficult to establish gaze-to-gaze contact with the wolves; therefore, wolves were less able to attend to an experimenter’s gestures for an extended duration (Miklósi et al., 2003; Gácsi et al., 2005; Virányi et al., 2006a). Dogs are inclined to look at our faces, and this inclination provides them with a broadened opportunity for learning about human gestures. Finally, we suggest that future researchers observing wolves in human-animal interactions should follow our lead. Wolves should receive extensive human contact individually beginning before the age of 10 days, for more than 20 hours a day, and should not be exposed to conspecifics early in life. Also, we believe that our insistence on early, extensive, and identical training in wolves and dogs make it likely that the differences we have observed are the result of species rather than experiential differences. Dogs as the Model of Socio-Cognitive Behavior in Humans According to Hare and Tomasello (2005), the major cause of behavioral differences between humans and apes is a basic change in the temperament. Selection against overt aggressive behavior, which had prevented the execution of complex cooperative actions in our ancestors, was the important change. This hypothesis is based on the observation that some animals, selected artificially for tameness (e.g., foxes, see Belyaev, 1978) show increased performance in understanding human cues (Hare & Tomasello, 2005). Although such changes may have contributed to the important behavioral differences between other primates and early human groups, we have argued that the evidence is too scant as yet to evaluate the role of temperament in human evolution (Miklósi & Topál, 2005; see also Byrne, 2005). We have proposed an alternative view that human evolution had an effect on not one, but many features of social behavior in a complex interacting way, during a stepwise process (Topál, Miklósi, et al., 2006; Byrne, 2005). These changes can be traced in the Human Behavior Complex (HBC) model which lists features of human social behavior that are assumed to have undergone alterations after the Pan-Homo split. Parallel changes in a different species could show that this model is viable. We think that dogs can provide a model because there seem to be a number of overlapping components in HBC and DBC, making dogs a fruitful comparative model for early social behavioral evolution in humans. We assume that divergence of dogs from wolves represents evolutionary steps taken in the same direction of increased sociability and cooperative and communicative abilities as observed in the case of the Pan-Homo clade. In our view, the comparative analysis of dogs and wolves should help scientists understand the first steps taken by our ancestors towards recent humans. References Ainsworth, M. D. S., & Wittig, B. A. (1969). Attachment and exploratory behavior of one-year olds in a strange situation. In B. M. Foss (Ed.), Determinants of infant behaviour (pp. 111-136). London: Meuthen. Altman, D. (1987). Social behavior patterns in three wolf packs at Tierpark Berlin. In H. Frank (Ed.), Man and wolf: Advances, issues and problems in captive wolf research (pp. 415-424). Dordrecht: W. Junk Publishers. Belyaev, D. K. (1978). Destabilizing selection as a factor in domestication. Journal of Heredity, 70, 301-308. Berryman, J. C., Howells, K., & Lloyd-Evans, M. (1985). Pet owner attitudes to pets and people: A psychological study. The Veterinary Record, 117, 659-661. Boitani, C., Francisci, F., & Ciucci, P. (1995). Population biology and ecology of feral dogs in central Italy. In J. Serpell (Ed.), The domestic dog: Its evolution, behavior and interactions with people (pp. 218-245). Cambridge: Cambridge University Press. Bradshaw, J. W. S., & Nott, H. M. R. (1995). Social and communication behavior of companion dogs. In J. Serpell (Ed.), The domestic dog: Its evolution, behavior and interactions with people (pp. 115-130). Cambridge: Cambridge University Press. Butler, J. R. A., du Toit, J. T., & Bingham, J. (2004). Free ranging domestic dogs (Canis familiaris) as predators and prey in rural Zimbabwe: Threats of competition and disease to large wild carnivores. Biological Conservation, 115, 369-378. Byrne, R. W. (2005). Animal evolution: Foxy friends. Current Biology, 15, R86-87. Cain, A. O. (1983). A study of pets in the family system. In A. H. Katcher & A. M. Beck (Eds.), New perspectives on our lives with companion animals (pp. 72-81). Philadelphia: University of Philadelphia Press. Cairns, R. B., & Werboff, J. (1967). Behavior development in the dog: An interspecific analysis. Science, 158, 1070-1072. Call, J., Bräuer, J., Kaminski, J., & Tomasello, M. (2003). Domestic dogs (Canis familiaris) are sensitive to the attentional state of humans. Journal of Comparative Psychology, 117, 257-263. Cohen, S. P. (2002). Can pets function as family members? Western Journal of Nursing Research, 24, 621-638. Coppinger, R. P., & Coppinger, L. (2001). Dogs. Chicago: University of Chicago Press. Csányi, V. (2000). The ‘human behavior complex’ and the compulsion of communication: Key factors in human evolution. Semiotica, 128, 45-60. Davey, G. (1981). Animal learning and conditioning. London: Macmillan Education. Davis, S. J. M., & Valla, F. R. (1978). Evidence for domestication of the dog 12,000 years ago in the Natufian of Israel. Nature, 276, 608–610. Donald, M. (1991). Origins of the modern mind. Cambridge, MA: Harvard University Press. Feddersen-Petersen, D. (1991). The ontogeny of social play and antagonistic behavior in selected canid species. Bonner Zoologische Beitraege, 42, 97-114. Feddersen-Petersen, D. (2000). Vocalization of European wolf (Canis lupus lupus L.) and various dog breeds (Canis lupus f. fam.). Archieves für Tierzüchtung (Dummerstorf), 43, 387-397. Fentress, J. C. (1967). Observations on the behavioral development of a hand-reared male timber wolf. American Zoologist, 7, 339-351. Finlayson, C. (2005). Biogeography and evolution of the genus Homo. Trends in Evolution and Ecology, 20, 457-463. Fox, M. W. (1970). Behavioral effects of rearing dogs with cats during the ‘critical period’ of socialisation. Behavior, 35, 273-280. Fox, M. W. (1971). Behavior of wolves, dogs and related canids. London: The Trinity Press. Fox, M. W. (1972). Socioecological implications of individual differences in wolf litters: A developmental and evolutionary perspective. Behavior, 45, 298-313. Fox, M. W. (1973). Social dynamics of three captive wolf packs. Behavior, 47, 290-301. Fox, M. W., & Stelzner, D. (1966). Approach/withdrawal variables in the development of social behavior in the dog. Animal Behavior, 14, 362-366. Frank, A. (1987). Man and wolf: Advances, issues and problems in captive wolf research. Dordrecht: W. Junk Publishers. Frank, H. (1980). Evolution of canine information processing under conditions of natural and artificial selection. Zeitschrift für Tierpsychologie, 5, 389-399. Frank, H., & Frank, M. G. (1982). Comparison of problem-solving performance in six-week-old wolves and dogs. Animal Behaviour, 30, 95-98. Frank, H., & Frank, M. G. (1985). Comparative manipulation-test performance in ten-week-old wolves (Canis lupus) and Alaskan malamutes (C. familiaris): A Piagetian interpretation. Journal of Comparative Psychology, 99, 266-274. Frank, H., Frank, M. G., Hasselbach, L. M., & Littleton, D. M. (1989). Motivation and insight in wolf (Canis lupus) and Alaskan malamute (Canis familiaris): Visual discrimination learning. Bulletin of Psychonomic Society, 27, 455-458. Freedman, D. G., King, J. A., & Elliot, O. (1961). Critical period in the social development of dogs. Science, 133, 1016–1017. Gácsi, M., Gyri, B., Miklósi, Á., Virányi, Zs., Kubinyi, E., Topál, J., et al. (2005). Species-specific differences and similarities in the behavior of hand raised dog and wolf puppies in social situations with humans. Developmental Psychobiology, 47, 111-122. Gácsi, M., Miklósi, Á., Varga, O., Topál, J., & Csányi, V. (2004). Are readers of our face readers of our minds? Dogs (Canis familiaris) show situation-dependent recognition of human’s attention. Animal Cognition, 7, 144-153. Gácsi, M., Topál, J., Miklósi, Á., Dóka, A., & Csányi, V. (2001). Attachment behavior of adult dogs (Canis familiaris) living at rescue centres: Forming new bonds. Journal of Comparative Psychology, 115, 423-431. Gomez, J. C. (1996). Ostensive behaviour in great apes: The role of eye contact. In A. E. Russon, S. T. Parker, & K. Bard (Eds.), Reaching into thought (pp. 131-151). Cambridge: Cambridge University Press. Gomez, J. C. (2005). Species comparative studies and cognitive development. Trends in Cognitive Sciences, 9, 118-125. Grzimek, B. (1942). Weitere Vergleichsversuche mit Wolf und Hund. Zeitschrift für vergleichende Physiologie, 5, 59–73. Gyri, B. (2004). Social attraction and communication in hand-raised wolves and dogs. Unpublished master’s thesis, Eötvös University, Budapest, Hungary. Hare, B., Brown, M., Williamson, C., & Tomasello, M. (2002). The domestication of cognition in dogs. Science, 298, 1634-1636. Hare, B., Plyusnina, I., Ignacio, N., Schepina, O., Stepika, A., Wrangham, R., et al. (2005). Social cognitive evolution in captive foxes is a correlated by-product of experimental domestication. Current Biology, 15, 226–230. Hare, B., & Tomasello, M. (2005). Human-like social skills in dogs? Trends in Cognitive Sciences, 9, 439-444. Hemmer, H. (1990). Domestication. The decline of environmental appreciation. Cambridge: Cambridge University Press. Klinghammer, E., & Goodmann, P. A. (1987). Socialization and management of wolves in captivity. In H. Frank (Ed.), Man and wolf: Advances, issues and problems in captive wolf research (pp. 31-60). Dordrecht: W. Junk Publishers. Kubinyi, E., Miklósi, Á., Topál, J., & Csányi, V. (2003). Social anticipation in dogs: A new form of social influence. Animal Cognition, 6, 57-64. Kubinyi, E., Topál, J., Miklósi, Á., & Csányi, V. (2003). Dogs learn from their owner via observation in a manipulation task. Journal of Comparative Psychology, 117, 156-165. Lakatos, G., Soproni, K., Dóka, A., & Miklósi, Á. (2006). A comparative approach of how dogs and human infants are able to utilize various forms of pointing gestures. Manuscript submitted for publication. Marston, L. C., Bennett, P. C., & Coleman, G. J. (2005). Factors affecting the formation of a canine-human bond. IWDBA Conference Proceedings, 132-138. Mech, L. D. (1970). The wolf: The ecology and behavior of an endangered species. New York: Natural History. Mech, L. D. (1999). Alpha status, dominance, and division of labor in wolf packs. Canadian Journal of Zoology, 77, 1196–1203. Mech, L. D., & Boitani, L. (Eds.). (2003). Wolves: Ecology, behavior and conservation. Chicago: University of Chicago Press. Meltzoff, A. N. (1996). The human infant as imitative generalist: A 20-year progress report on infant imitation with implications for comparative psychology. In I. C. M. Heyes & B. G. Galef (Eds.), Social learning in animals: The roots of culture (pp. 347-370). San Diego, CA: Academic Press. Miklósi, Á., Kubinyi, E., Topál, J., Gácsi, M., Virányi, Zs., & Csányi, V. (2003). A simple reason for a big difference: Wolves do not look back at humans, but dogs do. Current Biology, 13, 763-766. Miklósi Á., Polgárdi, R., Topál, J., & Csányi, V. (2000). Intentional behavior in dog-human communication: An experimental analysis of ‘showing’ behavior in the dog. Animal Cognition, 3, 159-166. Miklósi, Á., & Soproni, K. (2006). Comprehension of the human pointing gesture in animals: A comparative approach. Animal Cognition, 9, 81-93. Miklósi, Á., & Topál, J. (2005). Is there a simple recipe for how to make friends? Commentary. Trends in Cognitive Sciences, 9, 463-464. Miklósi, Á., Topál, J., & Csányi, V. (2004). Comparative social cognition: What can dogs teach us? Animal Behavior, 67, 995-1004. Mitchell, R.W. (2001). Americans’ talk to dogs during play: Similarities and differences with talk to infants. Research on Language and Social Interaction, 34, 182–210. Morey, D. F. (2006). Burying key evidence: The social bond between dogs and people. Journal of Archaeological Science, 33, 178-185. Morton, E. S. (1977). On the occurrence and significance of motivation-structural rules in some bird and mammal sounds. American Naturalist, 111, 855-869. Musiani, M., Mamo, C., Boitani, L., Callaghan, C., Gates, C.C., Mattei, L., et al. (2003). Wolf depredation trends and the use of fladry barriers to protect livestock in western North America. Conservation Biology, 17, 1538-1547. Naderi, Sz., Miklósi, Á., Dóka, A., & Csányi, V. (2001). Cooperative interactions between blind persons and their dog. Applied Animal Behavior Sciences, 74, 59-80. Nel, J. A. J. (1999). Social learning in canids: An ecological perspective. In H. O. Box & K. R. Gibson (Eds.), Mammalian social learning (pp. 259-277). Cambridge: Cambridge University Press. Nobis, G. (1979). Der älteste Haushund lebte vor 14000 Jahren. Umschau in Wissenschaft und Technik, 19, 610. Packard, J. M. (2003). Wolf behavior: Reproductive, social and intelligent. In L. D. Mech & L. Boitani (Eds.), Wolves: Behavior, ecology and conservation (pp. 35-65). Chicago: University of Chicago Press. Peters, R. (1978). Communication, cognitive mapping and strategy in wolves and hominids. In R. L. Hall & H. S. Sharp (Eds.), Wolf and man, evolution in parallel (pp. 95-107). New York: Academic Press. Plyusnina, I., Oskina, I., & Trut, L. (1991). An analysis of fear and aggression during early development of behavior in silver foxes (Vulpes vulpes). Applied Animal Behaviour Science, 32, 253–268. Pongrácz, P., Miklósi, Á., Kubinyi, E., Gurobi, K., Topál, J., & Csányi, V. (2001). Social learning in dogs: The effect of a human demonstrator on the performance of dogs in a detour task. Animal Behaviour, 62, 1109-1117. Pongrácz, P., Miklósi, Á., Kubinyi, E., Topál, J., & Csányi, V. (2003). Interaction between individual experience and social learning in dogs. Animal Behaviour, 65, 595-603. Pongrácz, P., Miklósi, Á., Timár-Geng, K., & Csányi, V. (2003). Preference for copying unambiguous demonstrations in dogs (Canis familiaris). Journal of Comparative Psychology, 117, 337-343. Pongrácz, P., Miklósi, Á., Timár-Geng, K., & Csányi, V. (2004). Verbal attention getting as a key factor in social learning between dog (Canis familiaris) and human. Journal of Comparative Psychology, 118, 375-383. Pongrácz, P., Molnár, Cs., Miklósi, Á., & Csányi, V. (2005). Human listeners classify dog barks in different situations. Journal of Comparative Psychology, 119, 136-144. Rabb, G. B., Woolpy, J. H., & Ginsburg, B. E. (1967). Social relationships in a group of captive wolves. American Zoologist, 7, 305-312. Reynolds, P.C. (1993). The complementation theory of language and tool use. In K. R. Gibson & T. Ingold (Eds.), Tool use, language and cognition in human evolution (pp. 407-428). Cambridge: Cambridge University Press. Savolainen, P., Zhang, Y., Ling, J., Lundeberg, J., & Leitner, T. (2002). Genetic evidence for an East Asian origin of domestic dogs. Science, 298, 610-613. Schassburger, R. M. (1993). Vocal communication in the timber wolf, Canis Lupus, Linnaeus: Structure, motivation, and ontogeny. Berlin: Paul Parey Scientific Publishers. Schenkel, R. (1967). Submission: Its features and function in the wolf and dog. American Zoologist, 7, 319-329. Scott, J. P., & Fuller, J. L. (1965). Genetics and the social behavior of the dog. Chicago: University of Chicago Press. Soproni, K., Miklósi, Á., Topál, J., & Csányi, V. (2002). Dogs’ (Canis familiaris) responsiveness to human pointing gestures. Journal of Comparative Psychology, 116, 27-34. Topál, J., Byrne, R., Miklósi, Á., & Csányi, V. (2006). Reproducing actions and action sequences: Do as I do in a dog. Animal Cognition, 9, 355-367. Topál, J. Gácsi, M., Miklósi, Á., Virányi, Zs., Kubinyi, E., & Csányi, V. (2005). The effect of domestication and socialization on attachment to human: A comparative study on hand reared wolves and differently socialized dog puppies. Animal Behavior, 70, 1367-1375. Topál, J., Kubinyi, E., Gácsi, M., & Miklósi, Á. (2005). Obeying social rules: A comparative study on dogs and humans. Journal of Cultural and Evolutionary Psychology, 3, 213-237. Topál, J., Miklósi, Á., & Csányi, V. (1997). Dog-human relationship affects problem solving behavior in the dog. Anthrozoös, 10, 214-224. Topál, J., Miklósi, Á., Dóka, A., & Csányi, V. (1998). Attachment behavior in dogs: A new application of the Ainsworth Strange Situation Test. Journal of Comparative Psychology, 112, 219-229. Topál, J., Miklósi, Á., Gácsi, M., Dóka, A., Pongrácz, P., Kubinyi, E., et al. (2006). An alternative functional model for the evolutionary study of human social behavior: The dog. Manuscript submitted for publication. Ujfalussy, D., Virányi, Zs., & Kubinyi, E. (2003). Individual and intensive hand-rearing of wolves: The method and its results in handling the wolves. ASAB Summer Meeting, Grünau, Austria. Újváry, D. (2004). Initiating communicative interactions with humans. Comparative analysis between dogs and wolves. Unpublished master’s thesis, Eötvös University, Budapest, Hungary. Vas, J., Topál, J., Gácsi, M., Miklósi, Á., & Csányi, V. (2005). A friend or an enemy? Dogs’ reaction to an unfamiliar person showing behavioural cues of threat and friendliness at different times. Applied Animal Behaviour Science, 94, 99-115. Vilá, C., Savolainen, P., Maldonado, J. E., Amorom, I. R., Rice, J. E., Honeycutt, R. L., et al. (1997). Multiple and ancient origins of the domestic dog. Science, 276, 1687-1689. Virányi, Zs., Gácsi, M., Kubinyi, E., Kurys, A., Miklósi, Á., & Csányi, V. (2002). Wolf-human interactions: Flight, approach and greeting behavior toward familiar and unfamiliar humans in hand-reared wolf pups (Canis lupus). Advances in Ethology (Supplements to Ethology), 37, 83. Virányi, Zs, Gácsi, M., Kubinyi, E., Topál, J., Belényi, B., Ujfalussy, D., et al. (2006a). Comprehension of human pointing gestures in young human-reared wolves (Canis lupus) and dogs (Canis familiaris). Manuscript submitted for publication. Virányi, Zs, Gácsi, M., Kubinyi, E., Topál, J., Belényi, B., Ujfalussy, D., et al. (2006b). Comparative analysis of ‘showing’ behavior of human-reared dogs and wolves. Unpublished manuscript, Eötvös University, Budapest, Hungary. Virányi, Zs., Topál, J., Gácsi, M., Miklósi, Á., & Csányi, V. (2004). Dogs respond appropriately to cues of humans’ attentional focus. Behavioural Processes, 66, 161-172. Yin, S. (2002). A new perspective on barking in dogs. Journal of Comparative Psychology, 119, 189-193. Zimen, E. (1987). Ontogeny of approach and flight behavior towards humans in wolves, poodles and wolf–poodle hybrids. In H. Frank (Ed.), Man and wolf: Advances, issues and problems in captive wolf research (pp. 275–292). Dordrecht: W. Junk Publishers. |






Enikő Kubinyi, Zsófia Virányi, and Ádám Miklósi, Department of Ethology, Eötvös University, Budapest, Hungary.
|