|
RESULTS
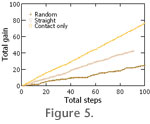 The simulations show, not surprisingly, that even a little knowledge of the outside chemical environment is tremendously advantageous (Table 1,
Figure 5,
Figure 6, and
Figure 7). In all cases, contact chemoreception alone produced an approximately threefold increase in the amount of resource gained, as compared to "blind wandering." The simulations show, not surprisingly, that even a little knowledge of the outside chemical environment is tremendously advantageous (Table 1,
Figure 5,
Figure 6, and
Figure 7). In all cases, contact chemoreception alone produced an approximately threefold increase in the amount of resource gained, as compared to "blind wandering."
For comparison, an additional run was made on the set 1 (fully occupied) map. In this run, the organism followed a simple straight path. This is the case labeled "straight" in
Table 1 and
Figure 5. Moving in a straight path increases the gain per step over random movements because it does not revisit already harvested nodes.
 What are more interesting are the relationships among contact and distant chemoreception and the patchiness of the map. In the case of a map where all nodes contained at least some resource, all runs with chemoreception produced exactly the same movement path (Figure 1.1), regardless of whether distant chemoreception was active. This was due to the organism never being out of direct contact with at least some resource. As discussed in
Koy and Plotnick (2007) paths on resource distributions of this type will avoid crossing themselves (i.e., show "phobotaxis"), because they do not move to already exploited areas. Because the distribution of resources is essentially the same in all directions, movement choice is governed by information of variation on very local scales. What are more interesting are the relationships among contact and distant chemoreception and the patchiness of the map. In the case of a map where all nodes contained at least some resource, all runs with chemoreception produced exactly the same movement path (Figure 1.1), regardless of whether distant chemoreception was active. This was due to the organism never being out of direct contact with at least some resource. As discussed in
Koy and Plotnick (2007) paths on resource distributions of this type will avoid crossing themselves (i.e., show "phobotaxis"), because they do not move to already exploited areas. Because the distribution of resources is essentially the same in all directions, movement choice is governed by information of variation on very local scales.
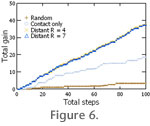
 When resources become patchier (Figure 1.2-1.3) the advantage of some form of distant chemoreception becomes clear. For the 20% occupied map, distant chemoreception produces a doubling of the average resource gained by step (Table 1,
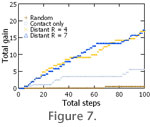
Figure 6). For the 5% occupied map, short range distant chemoreception (R = 4) nearly quadruples the gain rate, whereas longer range distant chemoreception (R = 7) increases it fivefold (Table 1,
Figure 7). When resources become patchier (Figure 1.2-1.3) the advantage of some form of distant chemoreception becomes clear. For the 20% occupied map, distant chemoreception produces a doubling of the average resource gained by step (Table 1,
Figure 6). For the 5% occupied map, short range distant chemoreception (R = 4) nearly quadruples the gain rate, whereas longer range distant chemoreception (R = 7) increases it fivefold (Table 1,
Figure 7).
Self-crossing is also infrequent on patchy resource maps. This is expected, since the simulated organisms will tend not to revisit regions that have already been harvested. When self-crossing does occur on the sparser maps, it is usually when the organism is out of sensory range of any resource and thus moves randomly in a small area.
 Many runs on the 20% map produced higher gain curves for short-range chemoreception than for long-range. Similarly, some runs on the 5% map also produced higher gains, at least for short periods, for short-range over long-range. Many runs on the 20% map produced higher gain curves for short-range chemoreception than for long-range. Similarly, some runs on the 5% map also produced higher gains, at least for short periods, for short-range over long-range.
This seeming conundrum can be understood by looking at the standard deviations of the mean resources gained at the end of the run. Every long-range run on the 20% map produced exactly the same path; because the detection-range was so long, it always located the "best" sites within this local range. Better sites in other parts of the map were missed.
On the other hand, the shorter-range chemoreception produced a greater randomness to the search. As a result, parts of the map that had slightly higher levels of resource were encountered. Some randomness in a search may lead to greater efficiency of resource acquisition. This result is consistent with theoretical studies of search strategies during foraging (Viswanathan et al. 1999), which suggest random movements increase search efficiency when the desired objective is not in direct sight.
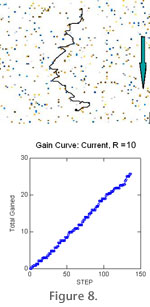
Introducing currents had only a minor effect on the search efficiency (Figure 8.2). The probability of detecting "upstream" sites was increased, but potentially better sites perpendicular to the current direction were missed.
|