|
|
|
Comparisons and discussionThe species described here is referred to the genus Lepidotes based on the following combination of characters that can be seen in the specimens: asymmetircal parietals; more than two anterior infraorbitals; a series of more than two suborbitals extending ventral to the orbit; close overlap between the opercle, interopercle, subopercle and propercle. Although all these features are present in the type species Lepidotes elvensis, none of these characters alone is unique to Lepidotes species, and indeed the genus is in urgent need of revision. There are well over 150 nominal species described from deposits ranging from the Rhaetic through Cenomanian. Almost any large Mesozoic fish showing thick rhomboid scales, thick skull bones and robust grasping and crushing teeth runs the risk of being identified as a species of Lepidotes. Many of these nominal species are based on very fragmentary material, poorly construed and must be considered as nomina dubia. Others are different parts of the same species, and some have been associated (see Woodward 1895). Yet others have been given specific names simply because of their geographic or stratigraphic locations. Clearly this situation only emphasises the need for a comprehensive review that must also include species referred to the genus Semionotus (another common species-rich taxon often confused with Lepidotes). An added dimension to this taxonomic uncertainty is the fact that the type species of the genus is Lepidotes elvensis (Blainville 1818) from the Lower Jurassic of Germany, France and England. This form differs considerably from many Upper Jurassic and Cretaceous species referred to the genus in the pattern of skull roof bones, numbers of extrascapulars, cheek bones, dentition and depth of mandibular symphysis (see Jain and Robinson 1963, Jain 1983). It is very likely therefore that many of the later species, including the taxon described here, will have to be referred to new genera. However, since this cannot be decided until much more revisionary work has been completed we keep this taxon in the genus, preferring this action to creating additional names that may not be justified. Despite the taxonomic uncertainty regarding the limits of the genus the taxon described here resembles strongly some of the Upper Jurassic and Lower Cretaceous species that show strongly tritoral dentitions such as the Upper Jurassic L. laevis Agassiz, 1837, L. toombsi Jain and Robinson, 1963, L. maximus Wagner, 1863, L. decoratus Wagner, 1863, or Lower Cretaceous L. degenhardti Branco, 1885, L. mantelli Agassiz, 1833, and L. souzai Woodward, 1908.
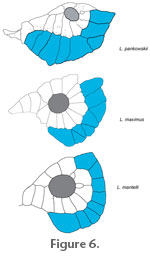
Among these tritoral species the cheekbone series varies, Jain (1983) noted that within the Lepidotes species that he considered (usually the better known Jurassic and Cretaceous species) there is a group containing species in which there are 2 – 6 suborbitals (Jain 1983 named these bones cheekplates) arranged in a single row. This is in contrast to a group containing species with 8 – 10 suborbitals (= cheekplates of Jain 1983). Lepidotes pankowskii falls into this latter group with at least eight suborbitals.
In all species studied so far, the suborbital series extends as far forward as the anterior level of the orbit. Lepidotes pankowskii differs in that this series extends well anterior to this level such that the anteriormost suborbital reaches close to the ethmoid region. Furthermore, the most anterior suborbitals in L. pankowskii are peculiarly sutured to the anterior infraorbitals. This feature is unique among the species of Lepidotes and among semionotiforms in general. Another feature noted by Jain (1983) and Woodward (1895) is the increase in the numbers of extrascapulars in later occuring species. Most of the non-tritoral species show a single pair of extrascapulars (e.g., L. semiserratus Agassiz, 1837, L. deccanensis Sykes 1851) while the tritoral species tend to show more (L. maximus – four pairs, L. mantelli – three or four pairs L. souzai – three pairs). Lepidotes pankowskii shows three pairs. A final observation noted by both Woodward (1895) and Jain (1983) is the fact that in later species the orbit decreases in relative size. The orbit of Lepidotes pankowskii is particularly small and may result in the apparent increased preorbital length and, perhaps the high number of anterior infraorbitals characteristic of this species. In sum L. pankowskii appears most closely similar to tritoral species such as L. mantelli and L. maximus in the dentition, disposition of the cheek plates and the number of extrascapulars. It remains distinct in the high number of anterior infraorbtials, the anterior extent and relationships of the suborbitals, and the small size of the orbit. The order Semionotiformes sensu Olsen and McCune (1991) is a monophyletic group including the lepisosteids, macrosemiids and semionotids. The monophyly of the Lepisosteidae and the Macrosemiidae are widely accepted, but the semionotids most probably represent a non-monophyletic assemblage including all semionotiforms that cannot be referred to one of the two monophyletic families (see López-Arbarello and Sferco (in press) for a brief historical overview). Nonetheless, the record of semionotid fishes in Africa is patchy and intriguing (López-Arbarello 2004, López-Arbarello et al. 2008). The oldest record in this continent is Semionotus capensis in the Early Jurassic Clarens Formation of South Africa (Woodward 1888), followed by Lepidotes congolensis in the Middle Jurassic Stanleyville Beds of the Lualaba Series in the Democratic Republic of Congo (Hussakof 1917, Saint-Seine 1955) and Lepidotes tendaguruensis in the Late Jurassic Upper Saurian Beds of Tendaguru (Arratia and Schultze 1999). With the exception of S. capensis, which is represented by numerous, relatively complete and rather well-preserved specimens, the other two species are only known from disarticulated bones or only a few and incomplete specimens respectively. Scales or teeth of Lepidotes have been reported from the Late Triassic-Early Jurassic Adigrat Sandstones, the Middle to early Late Jurassic Tiouraren Formation in Niger, and the Late Jurassic Mugher Mudstone Formation in Ethiopia (Moody and Sutcliffe 1991, Goodwin et al. 1999, Murray 2000, Rauhut and López-Arbarello 2009). Similarly, several isolated scales or teeth of Lepidotes have been mentioned throughout the Cretaceous (e.g., in the Albian Cocobeach Series of Gabon, the Albian Loia Beds and Albian to Early Cenomanian Bokungo Beds in the Democratic Republic of Congo, and the Hama Koussou Basin of Cameroon; more detailed information in Murray 2000), but articulated remains are very rare. Apart from Lepidotes pankowskii n. sp. described herein, complete and well-preserved specimens previously identified in Lepidotes and currently recognized as a different taxon, Pliodetes nigeriensis, are known from the Aptian of Gadoufaoua in Niger (Wenz 1999). Additional complete and well-preserved material identified as Lepidotes manni of probably Early Cretaceous age is reported from the Babouri-Figuil Basin in Cameroon, but this species is poorly understood and currently under study (Olga Otero, personal commun. 2010). Therefore, the only well-preserved material of semionotids is sparsely recorded from the Early Jurassic to the Cenomanian and represents quite different taxa. Even the species of Lepidotes, L. tendaguruensis and L. pankowskii are probably not closely related. Lepidotes tendaguruensis is strikingly similar to L. minor (ALA personal observations) from the Purbeck of England, which resembles Semionotus in several features (McCune 1986). As previously discussed, L. pankowskii is most probably closely related to the large tritoral forms known from the Late Jurassic and Early Cretaceous of central Europe. Lepidotes pankowskii furthermore represents the youngest confident record of a semionotid globally. Other Cenomanian or younger fossils identified in Lepidotes or simply referred to as semionotids consist of isolated bones or, more frequently scales or teeth. Although at least some of these fossils might actually represent semionotids, many of them might turn out to be lepisosteids after thorough revision. |
|