|
|
|
Systematic description
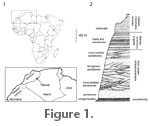
Subclass Neopterygii Regan, 1923 Diagnosis. Large Lepidotes reaching an estimated length of 1.6 m (based on body proportions of L. mantelli) in which the parietal is about half the length of the frontal; left parietal larger than the right; frontal-parietal suture interdigitate; three extrascapulars on either side of midline; orbit diameter small (12% of frontal-parietal midline length); complex suture between the nasal process of the premaxilla and the frontal; six circumorbitals with an additional small diamond-shaped circumorbital; four anterior infraorbitals; eight suborbitals with four lying in front of the orbit*; the most anterior suborbitals articulating with the anterior infraorbitals*; skull roof bones devoid of ganoine ornament. Lower jaw joint lying at a vertical level beneath the anterior edge of the orbit. [Asterisk denotes possible species autapomorphy.] Holotype. BMNH P.66856, partial head showing skull roof, cheek bones, preopercle, interopercle, part of the opercle and posttemporal, and partial lower jaw of left side, extrascapular series of right side, vomer and crushed braincase elements. Kem Kem Beds, southeastern Morocco (see below, Locality) Paratype. BMNH P.64126, skull roof and partial cheek of left side, vomer.
Etymology. Species named after Mr. Mark Pankowski of Rockville, Maryland, who kindly donated the holotype specimen to The Natural History Museum, London. We are grateful for his patronage of palaeontology.
The skull roofing bones conform in shape and pattern most closely to those in Lepidotes mantelli and L. maximus. Some of the measurements are shown in Table 1. As pointed out by Jain (1983, p. 34) some of the measurements of skull roofing bones are difficult because of extreme asymmetry between ipsilateral partners as well as the irregularity of suture lines. The midline suture length (frontals and parietals combined in P.64126 (the larger of the two specimens) is 235 mm. Assuming the proportions of the entire fish to be comparable with L. mantelli then this skull comes from a fish of about 1.6 m total length. The holotype would have been slightly smaller.
The parietals are asymmetrical (a common feature in Lepidotes species). In the two specimens described here the left parietal is approximately 110 percent larger than the right in area. As Jain (1983) noted in species of Lepidotes, it is usual that in the majority of individuals of any one species either the right or left element is larger, but that there may be in a few individuals showing opposite size relationships. Of course, with only two individuals we cannot be certain if this species favours larger left parietals. The dermopterotic reaches forward to an end level with the maximum forward extent of the parietal. Because the left parietal is larger than the right, there is a compensation in the dermopterotics sizes such that the right is slightly wider than the left. Many tiny pores run in a line close to the lateral edge of the dermopterotic marking the path of the otic sensory canal. All of the roofing bones show an ornament pattern of low vermiculating ridges. Ganoine appears to be absent from all skull bones. The absence of ganoine is unlike other species of Lepidotes. Most Jurassic species of the genus have an extensive covering of ganoine on the skull bones. Cretaceous species tend to have ganoine covering restricted to isolated tubercles on the roofing and cheek bones. In the species described here even that restricted ornament appears to be absent. We should add that weathering may have removed all traces; therefore we are cautious in claiming that this species is totally devoid of ganoine ornament. There are three extrascapulars on either side of the midline (only those of the right side are preseved in the holotype). The medial and lateralmost are both larger than the middle of the series, and in BMNH P.64126 the medial is larger than the lateralmost (in the holotype the medial is broken so the relative size is unknown). The medial extrascapular is sutured to the parietal, the lateralmost with the dermopterotic while the middle element sutures with both the parietal and the dermopterotic. In both specimens the dorsal tip of the posttemporal can be seen to project beneath the middle member of the extrascapular series and in BMNH P.64126 can be seen to reach the medial extrascapular. In BMNH P.64126 there are fragments of a median scale plus the first of the lateral scales.
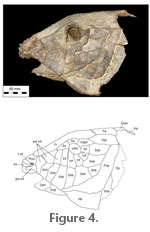
Remains of the right nasal can be seen in the holotype where it lies in situ. It is small and irregular in shape although the true shape is unknown because the anterolateral edge is broken. A line of pores crosses the nasal indicating the path of the supraorbital canal. The circumorbital and cheek bones are best preserved on the left side of the holotype (Figure 4). There is close agreement with the pattern of the remaining bones seen in BMNH P.64126. The orbit is very small compared with the length of the head, the maximum diameter being equal to about 12 percent mid-suture length of the skull roof. It appears to be considerably larger in most other species of Lepidotes examined here (e.g., in L. semiserratus this ratio is 30% while in L. mantelli it is 25%), except for L. roxoi (see Gallo and Brito 2004, figure 3) and probably L. latifrons (see reconstruction of the skull in Jain and Robinson 1963, figure 1). There are eight bones bordering the orbit. The two above the orbit are usually designated as supraorbitals, a large element at the posterodorsal position can be called the dermosphenotic since it carries the infraorbital sensory canal onto the roof where it presumably joins with the otic sensory canal, and five further elements border the posterior and anterior margins of the orbit. The two circumorbitals lying behind the eye are each equidimensional; those beneath the eye are deeper than wide. The supraorbitals and the dermosphenotic are sutured with the skull roofing bones. The elements behind and beneath the eye are marked with a prominent ridge along the orbital margin. This ridge may suggest the path of the main lateral line canal but it should be pointed out that there is a series of tiny pores scattered close to the distal margin of these bones that may mark the surface exits of the infraorbital sensory canal. Lying in front of the circumorbital series there are five more bones in the holotype, and these represent the anterior infraorbital and antorbital series (terminology of López-Arbarello and Sferco in press). In BMNH P.64126 only the most posterior of these is preserved. There are four anterior infraorbitals, two of them lie against the lateral edge of the frontal such that there is no gap between the skull roof and the anterior infraorbital series (cf., new semionotiform from Germany, López-Arbarello and Sferco in press). The next anterior infraorbital is a small triangular bone while the anteriormost is a curved element that lies above the nasal process of the premaxilla and is usually called the antorbital. Pores, presumably related to the infraorbital sensory canal, can be seen in all of the infraorbitals. They are most prominent in the anteriormost element as a line of closely spaced pores lying near the lateral edge of the antorbital. Between the dermosphenotic and the circumorbital behind the eye there is a tiny diamond-shaped bone that must be considered as part of the circumorbital series. It is found in both specimens where it is similarly developed and must be considered as a characteristic for the species. There is a series of eight suborbitals, seven of which are sutured to the bones of the infraorbital and antorbital series. At least three lie in front of the vertical level beneath the anterior margin of the orbit. The three most anterior suborbitals sutured with the four anterior infraorbitals, which is a unique feature among semionotiforms. The most posterior of the suborbitals is sutured to the dermosphenotic and the dermopterotic. A small suborbital lies at the posteroventral extent of the cheek series but it fails to reach the circumorbital series. Parts of the opercular bones are preserved on the left side of the holotype. The preopercle is prominent. It has an elongate vertical limb meeting a horizontal limb through an angle of about 120o. The horizontal limb reaches forward to the level of the jaw articulation. Scattered pores mark the path of the preopercular sensory canal. The interopercle is deep and appears tightly sutured to the whole horizontal limb of the preopercle. Behind this level parts of the subopercle and opercle can be seen, and all appear closely sutured to one another. The ascending process of the subopercle is broad, as is the case in Lepidotes maximus or L. laevis (contrary to L. mantelli or new semionotiform from Germany, López-Arbarello and Sferco in press). A small part of the left lower jaw is preserved in the holotype but little useful information can be gleaned except that the jaw joint is located at a vertical level beneath the anterior margin of the orbit. The angular is deep and shows a complex sinusoidal suture with the dentary (the anterior part of which is missing, so it is not possible to see the depth of the symphysis – a feature of some importance in distinguishing some Lepidotes species from others). Many small pores mark the path of the mandibular sensory canal running close to the ventral edge of the jaw. 1 The name Lepidotes pankowskii has appeared in Cavin (2010). This is a nomen nudum citation and does not alter the validlity of name published in the current paper. |
|