|
|
|
SYSTEMATICS
Order "CREODONTA" Cope, 1875 Diagnosis. Oxyaena differs from Dipsalidictis Matthew, 1915 in having a well-developed carnassial shearing dentition including a long postmetacrista on M1, and a long, often flaring, paralophid on M2. It also differs from Dipsalidictis in having lower molar trigonids longer than wide, in having reduced metaconids on lower molars, in having heavier premolars and in having a more anteroposteriorly compressed M2 that lacks a metacone. Type species. Oxyaena lupina Cope, 1874 Other species. Oxyaena forcipata Cope, 1874; Oxyaena gulo Matthew, 1915; Oxyaena pardalis Matthew, 1915; Oxyaena intermedia Denison, 1938; Oxyaena simpsoni Van Valen, 1966; Oxyaena woutersi (Lange-Badré and Godinot, 1982) Distribution. Early Eocene-Middle Eocene
Oxyaena woutersi (Lange-Badré and Godinot, 1982)
v. 1982 Arfia woutersi Lange-Badré and Godinot; p. 295-300, pl. 1, fig. 1; non pl. 1, figs. 2-4. Emended diagnosis. Smallest species of the genus. Morphologically intermediate between Dipsalidictis and Oxyaena. Shares with Dipsalidictis a trigonid wider than long on M1, lower molar cingulids and upper molar lingual cingula present but poorly developed and a postmetacrista on M1 less elongated than in other species of Oxyaena. It is closer to Dipsalidictis transiens Matthew, 1915 than to other species of that genus in the longer paracristid on the lower molars, a paraconid with a base more lingual on the lower molars, and a developed postmetacrista on M1. Shares with other species of Oxyaena a trigonid longer than wide on M2 and a talonid more robust on P3 and P4 (features also distinctive from Dipsalidictis transiens). Its closest Oxyaena species are Oxyaena sp. and Oxyaena gulo Matthew, 1915. It differs from later species of Oxyaena such as Oxyaena gulo by a trigonid much longer than wide on M2. Lectotype. M1 (or DP4?), IRNSB M1319 (L), figured as WL1147 by Lange-Badré and Godinot (1982; pl. I, fig. 1a-b).
MNHN QNY2-2527, left mandible with DP3, DP4 and M1, and alveoli of C1, P1 and DP2 MNHN QNY2-2569, right mandible with P4, M1 and trigonid of M2, and alveoli of C1, P1, P2 and P3 C1, MNHN QNY2-2561 (R), MNHN QNY2-2603 (L), MNHN QNY2-2584 (L), MNHN QNY2-2586 (L)
P4, MNHN QNY2-2551 (L), MNHN QNY2-2571 (R), MNHN QNY2-2574 (R), MNHN QNY2-2590 (R) M1, MNHN QNY2-2572 (R), MNHN QNY2-2573 (L), MNHN QNY2-2600 (L) P3, MNHN QNY2-2503 (L), MNHN QNY2-2549 (R), MNHN QNY2-2622 (R) P4, MNHN QNY2-2565 (L), MNHN QNY2-2595 (R) M1, MNHN QNY2-2579 (R) M2, MNHN QNY2-2524 (L)
Type locality. MP7, Dormaal (Belgium) Distribution. MP7, Dormaal (Belgium), Le Quesnoy (France).
Description
Premolars: As for all the teeth of Oxyaena woutersi, the enamel is slightly crenulated. Only P3 and P4 are known in Le Quesnoy. Both teeth have three roots and are morphologically similar. The P3 is distinctly smaller than the P4. The protocone is less developed on P3 than on P4: only a swelling is present lingually on P3 (no apex is present) while the protocone on P4 is well individualized and mesio-distally as long as the paracone. Contrary to the internal swelling on P3, the protocone of P4 is shifted mesially. On the two teeth, the lingual structure is located much lower than the paracone. The parastyle is absent on P3 and only poorly developed on P4. Both teeth present distally a very elongated and high postmetacrista separated from the paracone by a carnassial notch. The paracone is two-times higher than the postmetacrista. Its apex is tilted distally on the two teeth. P3 and P4 are surrounded by a tiny cingulum (slightly more pronounced on P4 than on P3). Molars: Only the M1 is known for this species. As seen on the fragmentary maxillary MNHN QNY2-2568, the M1 is similar in size to P4. The metacone and paracone are connate and are of equal development. The paracone is slightly higher than the metacone. The preparacrista is developed (but much smaller than the postmetacrista). It is mesio-labially oriented. It is separated from the paracone by a carnassial notch. The postmetacrista is very elongated and separated from the metacone by a deep carnassial notch. It is shifted distally (notably on MNHN QNY2-2573). The ectoflexus is very deep. The stylar shelf is very short labially. The protocone is transversally relatively narrow and is equal to the protocone of P4. Mesio-distally, it is shorter than the paracone and metacone, is shifted mesially and is also lower than the paracone and metacone (as for the premolars). The protocone is surrounded by pre- and postcingulum, but we do not know if they were linked lingually. The metaconule and paraconule are well developed. They seem to have a similar development. No internal crest is visible. The paraconule is linked to the parastyle by the preparaconule crista. The metacingulum reaches the top of the postmetacrista at its half-length. Excavations in the maxillary between the P4 and M1 and between the M1 and M2 are visible on MNHN QNY2-2568, probably to accommodate the tips of the lower molars (Gunnell and Gingerich 1991). Dentary. The two fragments of mandible found in Le Quesnoy show that the teeth were closely appressed. The mandible is slender and not very high. The symphysis extends below P3. It seems to have been very developed. Mandibular foramina are present below the P1 (near the tooth's crown) and the posterior root of P3. The coronoid crest is very pronounced and vertical. Lower dentition. Deciduous teeth: Two deciduous teeth (DP3 and DP4) are present in MNHN QNY2-2527. The teeth are very worn. Permanent teeth are visible inside the mandible (Figure 5). The DP3 and DP4 are compressed transversally and elongated mesio-distally, as is the typical morphology of deciduous teeth. The DP3 has a very low paraconid, probably poorly developed. The talonid is high, long (as long as the protoconid) and secant. The DP4 has a trigonid widely open lingually with a metaconid more distal than the protoconid. The paraconid is very slightly lingual. It seems to have been smaller than the metaconid. The talonid is as long as the trigonid and slightly narrower. The postfossid is much narrower than the trigonid. The talonid cusps are distally located, with a hypoconulid much more posterior than the hypoconid and entoconid. Premolars: Only the P1 is single-rooted. P2, P3 and P4 are double-rooted. The P3 is inserted obliquely in the dentary (the mesial part is lingual while the distal part is labial). Only P3 and P4 are known for this species. The teeth are morphologically similar but the P3 is distinctly smaller than the P4. The teeth are rather bulbous with a wide and short talonid. On P3 there is a fold in the mesio-lingual part of the protoconid but no distinct cuspate paraconid. The talonid is half as high as the protoconid. A carnassial notch on the cristid obliqua separates the protoconid from the hypoconid. The talonid has only one cusp, the hypoconid, which has a very wide base. The cristid obliqua is mesio-distal and curves into a posterior, ventro-lingually inclined, crest. Pre- and postcingulids are present. A tiny lingual cingulid is present along the talonid. A paraconid shelf is more developed on P4 than on P3 but is still not cuspate. The talonid of P4 is more elongated than on P3, but they are morphologically similar (e.g., orientation of the crest). The talonid is bulbous and wider than the protoconid. The cingulid on the lingual side of the talonid is more developed than on P3. A very tiny entoconid was possibly present. A very thin cingulid labially links the paraconid to the hypoconid. The mesial and distal parts of the tooth are very straight as on P3. Molars: The M1 has almost the same size as the P4. The trigonid is compressed mesio-distally. The bases of the paraconid and metaconid are in contact. The apices of the two cusps are slightly divergent. The apex of the metaconid is slightlty more posterior than the apex of the protoconid, and it is slightly tilted lingually. The prefossid is more closed than on deciduous premolars. The metaconid is only slightly higher than the paraconid. The protoconid part of the paracristid is directed mesio-distally; the paraconid part is much more transverse: the paraconid is not shifted mesially. Its base is slightly less salient lingually than the base of the metaconid. The protoconid is not much higher than the metaconid and paraconid and is not as sharp but is more robust. The paracristid and protocristid present a carnassial notch. A tiny cusp "e" is present on the mesio-lingual flank of the paraconid. The talonid is shorter than the trigonid, but, it is nearly as wide as the trigonid. The postfossid is narrow. The three cusps of the talonid are well developed. Two notches separate them. The separation is more pronounced between the hypoconulid and hypoconid than between the hypoconulid and entoconid. The hypoconulid is closer to the entoconid than to the hypoconid. The hypoconulid and entoconid have nearly the same height and are higher than the hypoconid. The base of the hypoconid is the largest. The cristid obliqua is disto-labially oriented. The part near the trigonid is worn, so it is impossible to know if a carnassial notch was present (a real possibility). The cristid obliqua runs along the trigonid distal wall toward the protocristid notch. The entocristid is oblique (distally shifted lingually) and closes the postfossid lingually. The curvature of the orientation of the entocristid and of the cristid obliqua renders the postfossid almost circular. The hypoflexid is deep. A pre-, ecto- and postcingulid link the paraconid to the hypoconulid. The M2 is higher, longer and sharper than the M1 and clearly represents the major carnassial tooth. Contrary to M1, the paraconid is slightly higher than the metaconid. The paracristid is less transverse than on M1: the paraconid is more shifted mesially. Its base is more extended lingually than that of the metaconid. A very tiny cusp "e" is present on the mesio-lingual flank of the paraconid. Moreover the protoconid is much taller than on M1 and secant. As on M1, the apex of the metaconid is slightly more distal than the apex of the protoconid. The metaconid is more reduced than on M1. The prefossid is more open than on M1. The paracristid and protocristid present a carnassial notch. The talonid of the M2 is more elongated mesio-distally than on M1 but it is distinctly narrower. The talonid cusps are also less developed than on the M1. Their apices are poorly defined. Numerous very tiny cuspules are present along the crests of the three cusps. Only a groove on the posterior wall of the basin separates the hypoconid from a hypoconulid part. This latter is the highest. The cristid obliqua and entocristid are mesio-distally oriented. Only a pre- and postcingulid are visible, but the base of the tooth is broken, so an ectocingulid may have been present. Discussion The fragment of mandible MNHN QNY2-2569 and the isolated molars MNHN QNY2-2579 and MNHN QNY2-2524 show clearly that the M1 is smaller than the M2 and that no M3 is present. On the maxillary fragment MNHN QNY2-2568 and on the isolated upper molars MNHN QNY2-2572 and MNHN QNY2-2573, the M1 has a well-developed postmetacrista. All these characters refer to the Oxyaenidae subfamily Oxyaeninae. The fossils differ from the genera Patriofelis Leidy, 1870 and Protopsalis Cope, 1880 by a less secant pattern of the molars and premolars: postmetacrista less developed on P4 and M1, protocone less reduced on P4 and M1, metaconid less reduced on lower molars. They are morphologically similar to Late Paleocene and Early Eocene genera of oxyaenines: Dipsalidictis and Oxyaena. The fossils found in Le Quesnoy have nearly the same size as Dipsalidictis platypus Matthew, 1915 and are distinctly smaller than all the other species known for the two genera in North America. By contrast to North American oxyaenines, our knowledge of European oxyaenines is very scarce. The best known European species is Oxyaena woutersi from the Belgian locality of Dormaal (MP7). O. woutersi is known by three teeth: two M1 and a P3 or P4 (Smith and Smith 2001). The teeth from Le Quesnoy have the same morphology as O. woutersi, but are slightly larger (12% longer for the P3) (Table 1). Our knowledge does not support species distinction based on this size difference between Le Quesnoy and Dormaal material. In oxyaenines species, the intraspecific variation in size can reach almost 10% (after Gunnell and Gingerich 1991, table 3). The M1 MNHN QNY2-2573 is more similar to IRNSB 1320 than to IRNSB 1319, which may be a DP4 (Smith and Smith 2001), or a M1 of Hyaenodontidae. They share a more pronounced ectoflexus, a more secant morphology, a more developed parastyle and a size larger than IRNSB 1319. The postmetacrista on M1 is slightly longer and more distally oriented in the material of Le Quesnoy. The lower premolar IRNSB 1318 was considered by Smith and Smith (2001) as a possible P3 or P4. Comparison of the material found in Le Quesnoy (notably the mandible MNHN QNY2-2569) shows that this specimen is a P3. The Le Quesnoy material is more complete than that of Dormaal and allows for a further discussion of the phylogenetic relationships of Oxyaena woutersi. Oxyaena and Dipsalidictis are poorly distinguished, and they often have been grouped together (Gunnell and Gingerich 1991). Dipsalidictis is the most primitive and appears during the Clarkforkian with Dipsalidictis krausei Gunnell and Gingerich, 1991 (Cf1-Cf3). Oxyaena, which appears during the Wasatchian with Oxyaena gulo (Wa2-Wa3), differs in having more secant upper and lower molars and more robust premolars. A difference between Dipsalidictis and Oxyaena is the absence of metacone on the M2 of the latter (Gunnell and Gingerich 1991). Unfortunately no M2 has been found in Le Quesnoy, and the presence/absence of the metacone on M2 can not be checked.
Some characters of the Le Quesnoy material are known in typical Oxyaena species. The P3 and P4 have a talonid enlarged labio-lingually as in Oxyaena. A maxillary excavation between the dental embrasures of M1 and M2 is typical of Oxyaena species. This character, which is derived in the oxyaenines, is linked to the strongly elevated M1 and M2 trigonid (Gunnell and Gingerich 1991). This maxillary excavation is distinctly present in MNHN QNY2-2568. The parastyle of P4 is smaller than in Dipsalidictis, similar in size to that of O. gulo. The postmetacrista of the M1 of O. woutersi is more elongated and shifted distally than in Dipsalidictis (but less than in O. gulo). O. woutersi is closer to O. gulo, the most primitive species of the genus, than to any other species of Oxyaena: they share a less-developed shearing dentition and narrower M2 talonid. On the other hand, some features of the upper teeth of the Le Quesnoy species recall the genus Dipsalidictis in having similar (and primitive) features: protocone on P4 more shifted mesially than on Oxyaena (this primitive morphology is also found in Tytthaena Gingerich, 1980); protocone of the M1 is less reduced than in O. gulo; deep ectoflexus on M1; parastyle developed on M1. The M1 trigonid is more compressed than in O. gulo. The labial cingulids of the M1 and M2 and pre- and postcingulum of the M1 are less developed than in O. gulo and other Oxyaena species. The presence of a small and poorly developed paraconid on P4 is known in some Dipsalidictis and the first species of Oxyaena, O. gulo. It is more developed in younger Oxyaena species. In general, Oxyaena woutersi has hypercarnivorous adaptations as known in Oxyaena (e.g., postmetacrista oblique and elongated, paracristid elongated on M2) however, some characters (e.g., paraconid lingually located and trigonid compressed in M1, which are primitive, recall also Dipsalidictis. The dental morphology of O. woutersi is intermediate between Dipsalidictis and Oxyaena, notably between D. transiens and O. gulo. Four species of Dipsalidictis are known: Dipsalidictis krausei (Cf1-Cf3), Dipsalidictis aequidens (Matthew, 1915) (Cf2-Cf3), Dipsalidictis platypus (Cf2-Wa0) and Dipsalidictis transiens (Wa0-Wa2). They are notably distinguished by their size. Except D. platypus, the Dipsalidictis species are larger than O. woutersi. O. woutersi differs from them notably by a longer paracristid on M2 and slightly longer postmetacrista on M1 which are derived features toward increased carnivory, which is characteristic of Oxyaena (see above). D. transiens has the most shearing teeth in the genus. Gunnell and Gingerich (1991) root Oxyaena in D. transiens. Dipsalidictis aequidens has a maxillary excavation between M1 and M2, as in Oxyaena species, including O. woutersi. Gunnell and Gingerich (1991) consider that this character is convergent with Oxyaena. D. aequidens has excessively larger and robust premolars (notably P4 with a well developed paraconid) and molars to be at the origin of Oxyaena, and of O. woutersi which has much more slender premolars and molars. This is in accordance with Gunnell and Gingerich (1991). The teeth of O. woutersi are much more secant than those of D. krausei. They differ notably in the paraconid of P4 more developed and in the protocone of the M1 and P4 less robust and mesio-distally elongated. These derived characters of O. woutersi are shared with D. platypus and D. transiens.
Our comparison suggests that the European O. woutersi continued to develop the shearing dentition which appeared in D. transiens, at the beginning of the Eocene. Among oxyaenines, O. woutersi is a structural intermediate between D. transiens and O. gulo. However, O. woutersi is distinguished by a M2 more secant than in O. gulo (known in one specimen only).
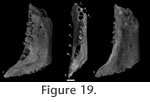
We retain the generic attribution of O. woutersi because its dentition is more specialized for shearing than Dipsalidictis (e.g., M2 paracristid enlarged), even if this concerns only the M2. Description of the Referred Postcranial Material from Le QuesnoyMeasurements: Table 4 Forelimb. MNHN QNY2-2526; left distal humerus. Only the distal part of the humerus is known. A slight torsion of the diaphysis is reflected by a somewhat posterior inclination of the supinator crest (origin of the elbow flexor muscle brachioradialis, and the forearm and digital extensors), which is not salient laterally. The distal part is transversally elongated. On medial and lateral views, the fragment appears almost flat. The trochlea is poorly projected anteriorly. The elbow joint is transversally elongated: the trochlea is well developed and the posterior trochlear groove is proximally deep and moderately concave. On the anterior side, the capitulum is less extended than the trochlea. The medial epicondyle is prominent and elongated medially. The entepicondylar foramen is very large. The medial lip of the trochlea is extended distally. In anterior view, there is a small and shallow radial fossa laterally to a round and large coronoid fossa. There is a hole in the coronoid fossa, although it does not seem to be the supratrochlear foramen, but rather the result of breakage. The pit for attachment of an ulnar collateral ligament is deep. The olecranon fossa is deep and wide. MNHN QNY2-2637; left scaphoid. The scaphoid is elongated mediolaterally (in dorsal view) and flat dorsoventrally (in mesial view). It is strongly convex dorsally. The scaphoid tubercule extends ventrally. The articular facet with the lunate is elongated proximodistally. In ventral view, the articular facets with the centrale (laterally) and trapezium (medially) are wide and square. The proximal facet is more oblique than of the proximal facet. The distal facets are almost flat, contrasting with the convexity of the proximal facet. Hindlimb. MNHN QNY2-2636; right calcaneum. The bone is short anteroposteriorly. The part posterior to the sustentacular facet is the longest; the anterior part to the facet is relatively short. The calcaneum is compressed mediolaterally, but dorsoventrally high. The tuber calcanei is wide, and worn laterally and medially. Posteriorly, there is a wide surface indicating a strong tendon insertion. The sustentacular facet (slightly concave) is more elongated anteroposteriorly than transversally. The facet is slightly inclined proximo-dorsally. The calcaneoastragalar facet (anteroposteriorly convex) is elongated anteroposteriorly, narrow and is directed dorso-posteriorly. The sustentacular facet and calcaneoastragalar facet are close to each other. There is also a very small calcaneofibular facet, posterior to the calcaneoastragalar facet. The peroneal process is broken distally, but, the large size of its base suggests that is was well developed. In anterior view the peroneal process is thin. Ventrally a surface for insertion is visible (for the calcaneocuboid ligament?). The articular facet with the cuboid is at an acute angle with the antero-posterior direction and slightly concave. In anterior view, the facet is somewhat rectangular. An articulation facet for the astragalar head is visible on the antero-dorsal part of the calcaneum. MNHN QNY2-2633, MNHN QNY2-2635; two right astragali. The astragali fit with the calcaneum in morphology and size. The astragalus is slightly elongated antero-posteriorly. The astragalar trochlea for the articulation with the distal tibia is shallowly grooved. The body is wide medio-laterally. The neck is narrower. It is robust and short antero-posteriorly. The head is slightly wider than the neck. It is convex anteriorly, but flattened dorso-ventrally. The astragalar foramen is well developed and weakly shifted posteriorly. In medial view, the medial part is lower than the lateral part. The sustentacular facet (convex) is broad and wide, but short antero-posteriorly and is inclined proximo-ventrally. The ectal facet is strongly concave. It is wide, but very thin antero-posteriorly. The sustentacular and ectal facets are well separated. Lateral to the ectal facet, there is a deep groove for the tendon of the plantaflexor muscles: flexor hallucis longus and peroneus longus. The head of the astragalus has a large and laterally extended navicular facet. MNHN QNY2-2638; cuboid. The cuboid is square except on the dorsal part. It is long proximodistally, but narrow transversally. The proximal facet, which articulates with the calcaneum, is convex and oblique. A triangular facet for the articulation with the astragalar head is present dorsally on the medial side. The articular facet with the navicular is extended anteroposteriorly, but short dorsoventrally. There is also a large articulation for the ectocuneiform. The plantar tubercule is well developed. It is laterally related to the distal articulation of the cuboid. The articular facet for the fourth and fifth metatarsals is transversally short and only slightly concave. DiscussionThe postcranial elements and the locomotion of the Oxyaenidae are well known. However, they have not been discussed in detail since the major work of Denison (1938). Only Rose (1990, 2001) and Gunnell and Gingerich (1991) discussed several points. A revision of the postcranium of the Oxyaenidae and a discussion of their locomotion are beyond the scope of this paper. The material of the Oxyaenidae from Le Quesnoy has been especially compared with the material housed in the collection of the AMNH and the University of Michigan. As explained previously, the Oxyaeninae are represented during the Paleocene and earliest Eocene by the genus Dipsalidictis, which is our main reference. The North American Oxyaena species are younger and more derived than the European O. woutersi. On the humerus (MNHN QNY2-2526) the medial trochlear rim is more salient distally than in Hyaenodontidae, and agrees with the Oxyaenidae. A second distal fragment of Oxyaenidae (MNHN QNY2-2634) differs from MNHN QNY2-2526 by a larger size and a more robust morphology, but, the trochleae are similar in shape: it belongs to Palaeonictis gigantea (see below). The distal fragment of humerus (MNHN QNY2-2526) is morphologically similar to AMNH 15857 and UM 69474 referred to D. platypus. Their sizes are also similar: as noted previously the two species have almost the same size. We refer MNHN QNY2-2526 to O. woutersi. The medial rim of the trochlea is not as extended distally as in O. forcipata and is more similar to Dipsalidictis platypus. The trochlea of MNHN QNY2-2526 is deeper posteriorly, less shallow than in D. platypus. The medial epicondyle (origin of muscle pronator teres and forearm and deep digital flexors), which is less well developed in terrestrial species than in arboreal and digging species (Taylor 1974; Argot 2003), is slightly more reduced than in Dipsalidictis. The radial fossa and the olecranon fossa are deeper than in Dipsalidictis. The lateral epicondylar crest, which is well developed in arboreal species (Heinrich and Rose 1997), is less developed in MNHN QNY2-2526 compared to Dipsalidictis. Moreover, the pit for attachment of the ulnar collateral ligament is slightly deeper. This ligament comes from the semilunar notch of the ulna, and it anchors the humerus and stabilizes the humeroulnar joint. The olecranon fossa and the great ulnar collateral attachment stabilize the elbow joint by reducing the axial rotation of the ulna. The supinator crest is less developed than in Dipsalidictis. The torsion of the diaphysis is less marked than in Dipsalidictis and is probably derived in comparison with the latter genus. All of these features are linked to a terrestrial locomotion that is slightly more pronounced in the forelimb of O. woutersi than in Dipsalidictis. The scaphoid is more elongated transversally than in Dipsalidictis. The dorsal facet is more developed in O. woutersi, thus limiting the flexibility of the manus. The calcaneum is similar in size to UM 66137 of D. platypus. It differs by a proximal articular facet that is extended proximo-distally and is more oblique (distolaterally oriented). Moreover the two articular facets with the astragalus are closer. The sustentacular facet is smaller. All these features are derived toward a less mobile ankle. A distinct fibular facet typical of the genus Oxyaena (Gunnell and Gingerich 1991) is also observable. The articular facet with the cuboid is flatter, elongated transversely and at an acute angle, which limits the movement between the two bones. This facet fits with the cuboid (MNHN QNY2-2638) found in Le Quesnoy. The proximal tuber extremity is worn. However, it may have been as large and developed as in Dipsalidictis. Its surface is more excavated than in Dipsalidictis. On the astragalus, the ectal facet is deeper than in D. platypus. However, the trochlea is similar. The articular facets for the calcaneum are more developed. The articular facets for the cuboid and navicular are more developed than in Dipsalidictis. The sustentacular facet contacts the navicular facet. It also extends closer to the ectal facet. This is a derived character toward a more terrestrial locomotion. The neck is shorter, more robust and broader. The fibular process is larger. All these features correspond to those of Oxyaena as noted by Gunnell and Gingerich (1991). They indicate a more robust ankle, with less flexibility. The astragali found in Le Quesnoy are less robust than in O. gulo: the body is lower, the trochlea is shallower and the ectal facet is less concave. In O. gulo, it limits the movement of the pes to a dorsoventral plane. The cuboid fits in structure and shape with the other fossils. The cuboid is shorter, but more robust than in Dipsalidictis (Gunnell and Gingerich 1991). As indicated above, its articulation with the astragalus is slightly more limited than in Dipsalidictis. On the whole, it appears that O. woutersi possessed more restricted articulations, which indicate movements more restricted to the parasagittal plane than in D. platypus. The trends here described are also found in the North America Oxyaena. O. woutersi thus possessed a more terrestrial locomotion than Dipsalidictis. Gunnell and Gingerich (1991) proposed that Oxyaena could be an ambulatory terrestrial predator, more terrestrial than Dispalidictis. Rose (2001) proposed the hypothesis that Oxyaena was semifossorial or scansorial, on the basis of an original combination of characters. Denison (1938) and Gunnell (1998) considered the Oxyaenidae as plantigrade. However, Rose (1990) proposed that the earliest Dipsalidictis were subdigitigrade. The smallness and robustness of the calcaneum (notably the tuber calcanei) is typical of plantigrade species (Ginsburg 1961). Thus, Oxyaena woutersi was probably plantigrade. One of the arguments used by Denison (1938) to propose an arboreal mode of life for Oxyaena was the development of the entepicondylar crest, here broken but surely developed. On this crest are inserted the pronation muscles. The supinator crest is present on the only available fragment of humerus, but reduced. This supports arboreal and/or scansorial habits, but less developed than in Dipsalidictis. The medial trochlear rim is more developed than in arboreal species. The presence of a delineated coronoid fossa proximal to the trochlea suggests usual use of highly flexed forelimb postures (Heinrich and Rose 1995, 1997). The coronoid fossa is found in arboreal and primitive carnivores and is linked to highly flexed forelimb postures. Its presence in O. woutersi is probably a primitive retention.The peroneal tubercle is also a primitive feature of arboreal carnivorous mammals. The peroneal musculature allows eversion, abduction and plantarflexion of the pes (Heinrich and Rose 1997). The plantarflexion role is predominant in terrestrial carnivorous mammals, which is not the case here. Plantarflexion is also limited by the high position of the astragalar foramen. Moreover a laterally extended peroneal process increases the leverage of the muscle peroneus longus (abductor of the hallux) (Argot 2004). The presence of a well-defined groove for the flexor hallucis longus tendon in the posterior side of the cuboid indicates that the flexor of the hallux was large. The hallux is somewhat divergent. On the astragalus, the differences in size of the medial and lateral trochlear crests indicate simultaneous inversion and abduction at the tibioastragalar joint during plantarflexion (Jenkins and McClearn 1984). The astragalus can slide anteriorly below the tibia: this increases the abduction and eversion at the tibioastragalar joint, and therefore allows the reversion of the hindfoot. However, these movements were rather limited. The presence of a developed lateral articular surface on the head indicates an usual eversion. Oxyaena woutersi, as Dispalidictis species, is clearly not cursorial. Many features can be related to an arboreal mode of life (e.g., divergent hallux, morphology of the astragalus). However, because the arboreal features are more developed in Dispalidictis species than in O. woutersi, we think that the arboreal features are primitive features. Moreover, the terrestrial features in O. woutersi are more developed than in Dispalidictis species. If we apply the methodology proposed by Morlo (1999) for body mass estimation of the "Creodonta" only on dental data, O. woutersi is estimated to have weighed approximately 7 kg, which is not in contradiction with an arboreal niche. However, O. woutersi was probably still capable of arboreal locomotion, but spending more time, possibly hunting, on the ground; this recent adaptative shift had not yet led to a marked terrestrial locomotion. However, several of the derived features of O. woutersi, compared to Dipsalidictis, are considered as adaptation to a more terrestrial locomotion. O. woutersi was probably a terrestrial ambulatory taxon.
Oxyaena sp.
Referred material from Meudon (France). M1, M-13-G (L) M1, MNHN ARP 5 (L) Referred material from Abbey Wood (England). M1, BMNH M31877 (L) M2, BMNH M13778 (R) Distribution. MP8+9, Meudon, Carrière des Montalets (Conglomérat de Meudon), Sinceny (France), Lessness Shell Bed, Blackheath Beds, Abbey Wood (England). DescriptionMeasurements: Table 5 Upper dentition. Molars: Two fragmentary M1 are known: M-13-G from Meudon (MP8+9) and BMNH M31877 from Abbey Wood (MP8+9). The trigon basin is missing on the M1. M-13-G is worn. However, its morphology is distinctly observable. The two specimens are morphologically similar. The paracone and metacone are connate but their apices are individualized. They are almost equal in development. The preparacrista is very small. It is more transversal than labio-lingually oriented. The postmetacrista is very elongated. It may have been separated from the metacone by a carnassial notch. It is shifted more distally than in O. woutersi. The ectoflexus is flat. The stylar shelf is very narrow. The paraconule and metaconule are present, and close to the paracone and metacone. The paraconule is linked to the preparacrista via the preparaconule crista. On the mesial part of the paraconule, a weak precingulum is visible. Lower dentition. Molars: One trigonid of a first molar is known from Meudon (MNHN ARP 5). UCMP 83754 from Sinceny is the only complete lower molar known in Europe for this species. It is slightly larger than MNHN ARP 5. However, the morphology of the two teeth is quite similar. They have a crenulated enamel. The trigonid is more elongated mesio-distally than in O. woutersi. The metaconid and paraconid are in contact. However their apices are divergent. The apex of the metaconid is slightly more distal than that of the protoconid. It is only very slightly titled lingually. The paraconid and metaconid are almost equal in height. The paracristid is distinctly less transverse than in O. woutersi and is also more elongated mesially. The paraconid projects mesially. The protoconid is not much higher than the paraconid and metaconid, but it is the highest cuspid. On M2, the lower carnassial tooth, the protoconid is much higher than the paraconid and metaconid. The paracristid and protocristid present a carnassial notch. The cusp "e" is present on the mesio-lingual flank of the paraconid but very slightly developed. The talonid is shorter than the trigonid. However, they have almost the same width. The postfossid is narrow and is surrounded by three well distinct cusps. The hypoconulid is more separated from the hypoconid than from the entoconid. These two cusps are close and have the same size. They are slightly higher than the hypoconid. The hypoconid has the largest base. The cristid obliqua is oblique (disto-labially oriented). No carnassial notch is present. The cristid obliqua extends on the trigonid wall towards the protocristid notch. The entocristid is oblique (distally shifted lingually) and closes lingually the postfossid. As in O. woutersi, thanks to the orientation of the entocristid and cristid obliqua, the postfossid seems to be almost circular. The hypoflexid is deep. A pre-, ecto- and postcingulid link the paraconid to the hypoconulid. The ectocingulid is well developed. The second lower molar here referred is a M2 from Abbey Wood (BMNH M13778). Only the paraconid is missing. By contrast to the previously described M1, the protoconid is tall and pointed. As on M1, the protoconid is distinctly higher than the metaconid (almost twice the height), and the metaconid is more reduced than on M1. Its apex is slightly distal to the protoconid producing an open prefossid. The talonid is shorter and narrower than the trigonid. By contrast to M1, the talonid is longer than wide, so the talonid appears to be elongated mesio-distally. The hypoconid is distinct but the entoconid is not as individualized. A slightly visible notch marks the separation between the entoconid and the hypoconulid. Their apices are poorly defined contrary to M1. The hypoconulid is more distal than the two other cuspids of the talonid. The entocristid is straight and aligned mesio-distally. The cristid obliqua is slightly oblique (distally shifted labially). The labial cingulids are well developed and are connected as on M1. DiscussionThe two teeth from Meudon MNHN ARP 5 (M1) and M-13-G (M1) (MP8+9, France) were not found during the same field campaign, but in the same facies ("Conglomérat de Meudon") and same locality. The other specimens here described and discussed are from Sinceny (MP8+9, France) and Abbey Wood (MP8+9, England). Van Valen (1965) referred the specimens from Abbey Wood to cf. Oxyaena sp. Gunnell and Gingerich (1991) consider MNHN ARP 5 and BMNH M31877 as representatives of a small Dipsalidictis species, BMNH M13778 as a large Dipsalidictis species, and UCMP 83754 as a Dipsalidictis or Oxyaena species. Hooker (1998) considers BMNH M13778 and BMNH 31877 as European representatives of O. gulo, and UCMP 83754 as Dipsalictis transiens. The teeth have well-developed secant structures (e.g., very long postmetacrista and paracristid, trigonid on molars longer than wide), which clearly permit to refer them to the Oxyaeninae. Oxyaena sp. has a paraconid more projected and a paracristid more elongated than in Dipsalidictis. The trigonid of the M1 of Oxyaena sp. is distinctly longer than wide. This feature is distinct for Oxyaena (see above). It also shares with other Oxyaena species a reduced metaconid, equal in height to the paraconid and not projected lingually. The metaconid is even smaller than in some specimens of O. gulo (e.g., UM 63670). The paraconid is broken on the M2. However, as noticed by Hooker (1998), the tooth belongs to the genus Oxyaena. The features shared with Oxyaena are the reduced metaconid compared to the protoconid, the talonid short and wide (but not as wide as the trigonid), the presence of well-developed postcingulid and ectocingulid. In M1, the postmetacrista is more shifted distally than in Dipsalidictis. The preparacrista is also more reduced and the postmetacrista is longer than in Dipsalidictis. The ectoflexus and stylar shelf are narrower than in Dipsalidictis. The derived morphology of the teeth of Oxyaena sp. permits to refer these teeth to the genus Oxyaena rather than to the primitive Dipsalidictis. UCMP 83754 (M1) from Sinceny is the only complete lower molar of an oxyaenine found in Europe, which is younger than the specimens of Le Quesnoy. UCMP 83754 is distinctly larger than the M1 of O. woutersi from Le Quesnoy (27% longer). UCMP 83754 also differs from the M1 of O. woutersi by a more secant morphology: the trigonid is longer than wide, the metaconid is more reduced (equal in height to the paraconid), the apex of the metaconid is projected less lingually and the paraconid is more shifted mesially. UCMP 83754 and MNHN ARP 5 (M1 from Meudon) are clearly derived toward more shearing dentition compared to the M1 of O. woutersi. The labial cingulid is more developed in the M1 of Oxyaena sp. from Sinceny than on the one of Le Quesnoy. The M1 of Oxyaena sp. is clearly more derived compared to the M1 of O. woutersi because all its features (e.g., paraconid more projected mesially, metaconid reduced, ectocingulid more developed) are derived within the Oxyaeninae. BMNH M13778 is the only M2 of Oxyaena known for fossiliferous localities younger than Le Quesnoy in Europe. Compared to MNHN QNY2-2524 of O. woutersi, BMNH M13778 of Oxyaena sp. is distinctly larger (26% wider). BMNH M13778 has also more developed labial cingulid, reduced metaconid, and talonid shorter and wider. These are derived trends observed in the North American Oxyaeninae. The upper molars of Oxyaena sp. from Meudon and Abbey Wood are distinctly bigger than those of O. woutersi (19% longer). Oxyaena sp. also differs by a more elongated and distally shifted postmetacrista, narrower stylar shelf, narrow ectoflexus, and very short parastyle. As for the lower molar of Oxyaena sp., all these features are derived within the Oxyaeninae. We choose to group all the specimens in Oxyaena sp. despite their different origin, because they share similar morphology and derived features, notably the mesial location of the paraconid on molars, and the elongation and distal location of the postmetacrista. Their derived features clearly distinguish the fossils of Oxyaena sp. from O. woutersi. The derived features are not restricted to Oxyaena sp. However they support a distinction between Oxyaena sp. from Meudon, Abbey Wood and Sinceny, and O. woutersi from Dormaal and Le Quesnoy. Oxyaena sp. is also larger than O. woutersi. Its size is closer to the oldest specimens of O. gulo. We prefer presently not to erect a new species because the specimens of Oxyaena sp. are fragmentary and few. All the derived features of Oxyaena sp. (e.g., trigonid longer than wide on M1, paraconid as high as the metaconid, cingulids developed, postmetacrista long and shifted mesially) are observed in O. gulo. However, UCMP 83754 shares with the M1 of Le Quesnoy an unusual talonid, which distinguishes Oxyaena sp. from O. gulo.
The combination of oblique entocristid and cristid obliqua, and the more distally projected hypoconulid, gives a rounded morphology to the talonid of the European Oxyaena. In the North American Oxynaena, the hypoconulid is more mesially located, and the talonid of the North American Oxyaena is wider and more squared. The M1 from Sinceny appears to be morphologically closer to the European O. woutersi than to the North American O. gulo. This feature permits to distinguish Oxyaena sp. from O. gulo. The morphology of the talonid is the only present distinctive feature of the two European Oxyaena compared to O. gulo. The two European Oxyaena species also share the absence of an individualized entoconid. In O. gulo, as in all younger North American Oxyaena, the entoconid is separated from the hypoconulid by a deeper notch than in Dipsalidictis. The close entoconid/hypoconulid and the morphology of the talonid are primitive features within the Oxyaenidae: they are notably present in the oldest Oxyaenidae Tytthaena lichna (Rose, 1981) and in Dipsalidictis. The morphology of the European Oxyaena is primitive compared to O. gulo. Oxyaena woutersi and Oxyaena sp. represent probably an endemic clade of Oxyaena characterized by the retention of primitive features (Figure 9). Oxyaena sp. is more derived than O. woutersi notably in having an M1 more secant, which suggests that the evolution toward secant molars is convergent with the evolution observed in the North American Oxyaena. However, we prefer inclusion of the European oxyaenines within the single genus Oxyaena. Conclusion on the European OxyaeninaeThe Oxyaeninae were poorly known before the discovery of Le Quesnoy material, because fossil oxyaenids are rare in Europe. The fossils from Le Quesnoy add further data on the dentition of the earliest European representatives of Oxyaenidae. Comparisons suggest that European Oxyaena represent a lineage different from North American Oxyaena. The two lineages from Europe and North America are derived from the same Dipsalidictis species: D. transiens. The difference between the North American and European Oxyaena lineages is based on the distinctive morphology of the talonid of M1, which is more primitive in the European Oxyaena. However, the European Oxyaena species present other features related to the development of a specialized secant dentition. These features are similar to what is known in the North American Oxyaena, notably in the oldest species, O. gulo. Because of the more primitive morphology of O. woutersi compared to Oxyaena sp. from Meudon, Sinceny and Abbey Wood (e.g., M1 wider than long in O. woutersi), we think that the development of secant structures in the European lineage is convergent with North American Oxyaena. Aside from the development of the secant structures, European Oxyaena are characterized by an increase in the size; this is also observed for the North American Oxyaena (see Gunnell and Gingerich 1991). The larger size is recorded at the beginning of the MP8+9 with the fossils from Meudon, Abbey Wood and Sinceny. This species also corresponds to the last occurrence of Oxyaeninae in Europe. Oxyaena woutersi and Oxyaena sp. provide evidence for an endemic evolution of the Oxyaeninae in Europe (Figure 10). This lineage supposes a single dispersal event of Oxyaeninae in Europe from North America, right near the Paleocene-Eocene boundary.
Subfamily AMBLOCTONINAE Cope, 1877 Diagnosis. Palaeonictis differs from Dipsalodon Jepsen, 1930 in having M2, much smaller than M1, in having a relatively smaller metaconid on M2, with a more open trigonid, and in having a less well-developed talonid on P4 that lacks an entoconid cusp. Type species. Palaeonictis gigantea de Blainville, 1842 Other species. Palaeonictis occidentalis Osborn and Wortman, 1892; Palaeonictis peloria Rose, 1981; Palaeonictis wingi Chester et al., 2010 Distribution. Late Paleocene-Early Eocene
Palaeonictis gigantea
de Blainville,
1842
v*1966 Oxyaena (?) casieri Quinet, p. 35, pl 5, figs. 4-6 v 1987 Dormaalodon woutersi Lange-Badré, p. 831, fig. 1 Syntypes.
I3, MNHN ARP 53 (L) P2, MNHN ARP 54 (L) Referred material from Le Quesnoy. C1, MNHN QNY2-2562 (L), MNHN QNY2-2588 (R)
P2, QNY1-1309 (L), MNHN QNY2-2589 (L), MNHN QNY2-2599 (L) P3, MNHN QNY2-2555 (L), MNHN QNY2-2557 (L), MNHN QNY2-2559 (R), MNHN QNY2-2566 (R), MNHN QNY2-2592 (L) P4, MNHN QNY2-2558 (R), MNHN QNY2-2563 (R), MNHN QNY2-2564 (R), MNHN QNY2-2587 (L) M1, MNHN QNY2-2567 (R), MNHN QNY2-2575 (R) C1, MNHN QNY2-2580 (R), MNHN QNY2-2621 (L), MNHN QNY2-2626 (R) P2, MNHN QNY2-2597 (L)
P4, MNHN QNY2-2556 (L), MNHN QNY2-2560 (L) M1, MNHN QNY2-2550 (L), MNHN QNY2-2552 (R) M2, MNHN QNY2-2513 (R), MNHN QNY2-2576 (L), MNHN QNY2-2577 (R), MNHN QNY2-2578 (L) Referred postcranial material from Le Quesnoy. MNHN QNY2-2634, right distal humerus; MNHN QNY2-2639, right radius; MNHN QNY2-2604, left astragalus; MNHN QNY2-2605, left astragalus. Referred material from Meudon. M1, MNHN ARP 6 (L)
P4, MNHN L-PY-60 (R) Type-locality. Muirancourt, Argiles à Lignites du Soissonnais (MP7) for MNHN ARP 52; unknown locality, Argiles à Lignites du Soissonnais (MP7) for MNHN ARP 53 and MNHN ARP 54. Distribution. MP7, Muirancourt, Argiles à Lignites du Soissonnais, Le Quesnoy, Pourcy, Houdancourt (France), and Dormaal (Belgium); MP8+9, Meudon (France). Description of the dental material from Le QuesnoyMeasurements: Table 6
Canines: Two specimens are known. The tooth is distinctly bigger than in O. woutersi. They are oval in horizontal section. The root is very large. The cingulum is poorly developed. However, it rises a little in the lingual part of the tooth and continues as a crest. The tooth is straighter than the lower canines, and the apex is slightly more shifted lingually. There is a second carina in the disto-lingual face of the tooth. A wear facet is present in the mesio-labial part of the tooth. Premolars: The P2, P3 and P4 are known. Their size increases with the distal rank. The P2 and P3 are two-rooted; the P4 has three roots. The premolars share the same pattern. The paracone is large and bulbous. Its apex is tilted distally (also slightly lingually on P2 and P3). The parastyle is only marked by a fold in the mesio-lingual part of the tooth on P2 and P3 - it is slightly more developed on P3, and much more developed and indvidualized on P4, where it is located mesial to the paracone. It reaches one-third the size of the paracone. The postmetacrista is very short and bulbous on all premolars. It reaches half the height of the paracone on P4. The protocone is marked on P2 and P3 by a little fold. On P4, the protocone is well developed and functional. It is lower than the paracone, as seen on M1. The protocone is slightly shifted mesially. A thin cingulum surrounds the teeth. It elevates distally to the postmetacrista and on P4 also mesially to the paracone. Molars: Only M1 is known in Le Quesnoy. The paracone and metacone are robust and almost fully separated. The stylar shelf is absent. Only a little cingulum links labially the parastyle to the metastyle. The preparacrista is very reduced. Only a small enamel fold at the half height of the paracone represents the parastyle. The postmetacrista is more developed than the preparacrista, but very poorly elongated compared to what is seen in Oxyaena. The postmetacrista is almost aligned with the paracone and metacone. It is more than half the size of the metacone. The metacone is slightly higher than the paracone and also slightly larger. The protocone is lower than the labial part of the tooth. It is wider labio-lingually than mesio-distally. The metaconule and paraconule are present and well developed. They are connected respectively to the postmetacrista and preparacrista. No internal crest is present. The protocone is very weakly shifted mesially. The pre- and postcingulum are present but are poorly developed; they are linked lingually. Lower dentition. Canines: Three lower canines are known. These teeth are more curved and slightly larger than the upper canines. Two crests are present in the lingual part and in the disto-labial part. On MNHN QNY2-2621 and MNHN QNY2-2580, a strong wear facet is visible in disto-labial position. This facet results from the occlusion with the upper canine. Another wear facet is visible on the mesio-labial part of the tooth, as the result from the occlusion with the I3. Premolars: As for upper premolars, the first premolar is unknown. The size of lower premolars increases with the distal rank. The teeth are two-rooted. The protoconid, which is very bulbous, constitutes the main structure of the teeth. The protoconid is tilted posteriorly and labially on P3 and P4 (probably also on P2). The paraconid is marked by a fold in the mesio-lingual facet of the protoconid in P2 and P3. On P4 it is well individualized. It is less high than the talonid. The talonid is short and bulbous. It reaches half the size of the protoconid. It is constituted by a single cusp (hypoconid) distally shifted lingually. It is separated from the protoconid by a carnassial notch. It is more shifted on P3 and P4 than on P2. The talonid on P4 is variable. The talonid extends up distally in MNHN QNY2-2556 and extends down in MNHN QNY2-2560. On P4, an entoconid is present lingually. A cingulid surrounds the tooth on P2 and P3. Only a labial cingulid runs from the labial facet of the paraconid to the hypoconid on P4. Molars: The M1 and M2 are known. The teeth are two-rooted. The posterior root of M2 is elongated mesio-distally. Contrary to O. woutersi, the M1 is wider and higher than the M2. and it represents the carnassial tooth. In M1, the paraconid and metaconid have nearly the same height. The apices of the two cusps are very divergent. The prefossid is closed lingually by a small contact of the bases of paraconid and metaconid. The metaconid apex is distinctly more distal than the protoconid apex. The paraconid apex is much less lingual than its base. The paracristid is very short and almost transverse. These characteristics explain the shortness of the paracristid, almost only constituted by the protoconid crest. The paracristid and protocristid present a carnassial notch. There is an accessory cusp "e" in the mesio-lingual face of the paraconid. The protoconid is also very bulbous with a well-developed base. It is higher (but not very much) than the other cusps. The talonid is narrower and shorter than the trigonid. The talonid bears three well developed and separated cusps. All the cusps are high, the hypoconid being the highest. The hypoconulid is more distal than the two other cusps. Its distal facet is almost straight. The hypoconid is slightly more mesial than the entoconid. The cristid obliqua is very oblique (distally shifted labially). A carnassial notch is present near the trigonid. The cristid obliqua rises very high on the distal facet of the trigonid toward the apex of the metaconid. The entocristid is aligned with the mesio-distal axis. The pre-, ecto- and postcingulid are present. Another cingulid runs lingually from the cusp "e" to the base of metaconid. M2 looks like M1. The apex of the paraconid is less lingual than its base, and the mesio-lingual facet is curved. The crest of the paraconid is less reduced than on M1 but is also transverse. The metaconid's apex is more distal than that of the protoconid. The metaconid is also distinctly smaller than the paraconid. The prefossid is opened: the base of the protoconid separates the paraconid and the metaconid. The protoconid is bulbous and not much higher than the paraconid. The paracristid and protocristid present a carnassial notch. The accessory cusp "e" in the mesio-lingual flank of the paraconid is less individualized than on M1. The principal difference between the M1 and M2 is that the talonid is narrower, shorter and smaller than the trigonid on M2. Only the hypoconulid and hypoconid are well developed on the talonid of M2, and the two cusps are distally placed. They tend to fuse in some teeth (MNHN QNY2-2513). On some specimens, where the fusion is the less pronounced, an entoconid is visible. The cristid obliqua is oblique (distally shifted labially) and, on some teeth, it joins the apex of the metaconid. A carnassial notch is present. The distal facet of the talonid is very straight. The cingulids are similar to what is seen on M1 (pre-, ecto- and postcingulid and labial cingulid between the cusp "e" and base of the metaconid).
Description of the Material from the Syntypes Upper dentition. MNHN ARP 53 and MNHN ARP 54 have not been described by Blainville (1842). These fossils may have been added posteriorly to the syntypes of P. gigantea. Their colours and appearance agree with fossils from the Argiles à Lignites. However, the exact locality remains unknown for them. Incisive: MNHN ARP 53 represents an I3. This tooth is characterized by a strong occlusal facet in the disto-labial part of the tooth, which is caused by the occlusion with the lower canine as discussed above. MNHN ARP 53 does not differ from the fossils from Le Quesnoy, except by a slightly larger size. Premolars: MNHN ARP 54 is a left fragment of maxillary with P2. MNHN ARP 54 is also indistinguishable from MNHN QNY2-2597 of Le Quesnoy. Lower dentition. Premolars and molars: MNHN ARP 52 corresponds to a fragmentary mandible. This fossil is the type of P. gigantea and was previously described by de Blainville (1842). Since this work, the fossil has been broken up. No foramen is visible on this fragment of mandible. The mandible is deep and robust as in all Palaeonictis. The lingual part is more robust than the labial part. The teeth are inserted in a parabolic way. The first tooth of the specimen MNHN ARP 52 is a P3. The morphology of the tooth (e.g., presence of a small paraconid) agrees with the P3 of P. occidentalis and P3 found in Le Quesnoy. The P3 is the sole complete remaining tooth. The P3 is robust and, as all the other teeth, has a crenulated enamel. The protoconid is bulbous. There is a paraconid mesio-lingually located. The paraconid is separated from the protoconid by a distinct notch and is one-third the size of the protoconid. The talonid is also separated from the protoconid by a notch. The talonid is half the size of the protoconid in height and length. However, the talonid is wider than the protoconid, notably lingually. The talonid has only one cusp. It is oblique (distally shifted lingually). Its distal part is higher than the mesial part. A cingulid is present lingually. Labially, only precingulid and postcingulid are present. They are not linked by an ectocingulid. On the lingual part of the talonid, two very small accessory cusps are present. They are located at the usual place of the entoconid on the P4. The P3 preserved on MNHN ARP 52 only differs from the P3 found in Le Quesnoy (MNHN QNY2-2591) by more developed paraconid and cingulids. The talonid of the P4 is similar in structure to the talonid of P3. The P4 of MNHN ARP 52 differs also from the P4 found in Le Quesnoy by a more pronounced cingulid around the talonid and less pronounced entoconid in the lingual part of the talonid. Our observations of the specimens of Palaeonictis occidentalis housed in the collections at the University of Michigan document that these characters are highly variable in this species. However, this reflects a tendency toward the reduction of the entoconid: in Dipsalodon, the entoconid is more elevated, while it is absent in the younger Ambloctonus Cope, 1875. As for the P4, only the talonid is available for the M1. The talonid of the M1 has the same morphology as the fossils from Le Quesnoy. The derived genus Ambloctonus is characterized by an entoconid well isolated and low. This feature is not visible in MNHN ARP 52, as usually observed on M1 of Palaeonictis.
Description of the Material from Meudon Lower dentition. Molar: MNHN ARP 6 is a trigonid of first lower molar. MNHN ARP 6 is characterized by ambloctonine features such as the metaconid distal to the protoconid and the trigonid robust. The strong specialization of the basic trigonid structure (metaconid well distal, very short and labial paraconid) permits reference of this tooth to the genus Palaeonictis rather than Dipsalodon. Ambloctonus differs in the cusps of the trigonid, which are nearly similar in size. MNHN ARP 6 remains primitive compared to Ambloctonus. MNHN ARP 6 very slightly differs from the M1 found in Le Quesnoy by a metaconid more distal, an apex less projected lingually, a paraconid less lingual, a more pronounced labial cingulid and a prefossid more opened. These correspond to trends observed in North American Palaeonictis toward the unusual dentition of Ambloctonus. MNHN ARP 6 from Meudon is slightly more derived than the specimens from Le Quesnoy. However, MNHN ARP 6 is not distinctly larger than the M1 from Le Quesnoy of Palaeonictis gigantea. As we do not know the entire dentition of this taxon and its variability, we refer MNHN ARP 6 to P. gigantea.
Description of the Material from Pourcy Lower dentition. Premolar: The P4 MNHN L-Py-60 from Pourcy is slightly larger than the fossils from Le Quesnoy, but slightly smaller than the P4 of MNHN ARP 52 from "Argiles à lignites". A broken part suggests an individualized paraconid. MNHN L-Py-60 is referred by Gunnell and Gingerich (1991) to Dipsalidictis. By contrast this fossil is considered here as representative of Palaeonictis. MNHN L-Py-60 differs from Dipsalidictis species in being much more robust and in possessing a talonid much wider than the trigonid. It differs from Oxyaena by a talonid expanded lingually. In Oxyaena the talonid is widened lingually but also labially. Moreover, the tooth is more similar in size and structure to the P4 of Palaeonictis found in Le Quesnoy, than to those of Oxyaena. It notably presents some small accessory cusps on the lingual part of the talonid. No similar cusps are visible on the P4 of Oxyaena and Dipsalidictis. MNHN L-Py-60 is similar to MNHN ARP 52 in having a shorter talonid than in the P4 from Le Quesnoy. The cingulids are less developed on MNHN L-Py-60 than on MNHN ARP 52 and recalls more the fossils from Le Quesnoy. Affinities of DormaalodonIn Dormaal, Lange-Badré (1986, 1987) has described a new genus, Dormaalodon Lange-Badré, 1987 from a very peculiar lower molar. This genus has been only found in Dormaal. Lange-Badré (1987) described the lower molar (IRNSB M 1474) as the holotype and only known specimen of the ambloctonine Dormaalodon woutersi. This tooth presents some puzzling characters: metaconid posteriorly located, separated from the protoconid and integrated to the talonid; absence of entoconid; cingulid well developed, which reaches lingually the metaconid (Figure 16). The molar is considered by Lange-Badré (1987) as a M1 because of the presence of a posterior interdental facet indicating a contact with a posterior molar, the M2. Our observations do not confirm the occurrence of this facet, and there are no features supporting IRNSB M 1474 as a M1. Comparison with the M2 of Palaeonictis gigantea found in Le Quesnoy shows that IRNSB M 1474 is slightly shorter, as for the P4 of Dormaal. IRNSB M 1474 presents an important wear: notably the mesio-labial facet of the paraconid and protoconid (wear facet 2), distal facet of the trigonid (wear facet 1) and mesio-labial facet of the hypoconid (wear facet 3), which are very marked; for instance, the hypoconid is even partially truncated (Figure 16). These facets are similar to those observed on the M2 from Le Quesnoy. Wear facet 3 is caused by the occlusion with the paracone of the M2. We can note the high development of the cristid obliqua, which runs along the distal facet of the trigonid toward the carnassial notch of the protocristid and apex of metaconid. The cristid obliqua visible on the M1 is less developed than on M2 but presents an almost similar morphology. Wear facet 1 is caused by the long and robust paracrista of the M2. Wear facet 2 is caused by the poorly elongated but high and robust postmetacrista and metacone of the M1. Wear facet 4 of the M1 is caused by the occlusion with the metacone, which has almost the same size as the paracone on M1. This facet is not present on the M2 from Le Quesnoy nor is it present on the only known molar of Dormaalodon due to the reduction of the metacone of the M2 in the Oxyaenidae. Contrary to IRNSB M 1474, the protocristid is not broken on the M2 found in Le Quesnoy. The metaconid is linked to the protoconid and not included in the talonid. The cristid obliqua is well developed and, on some M2, it reaches the apex of the metaconid through the carnassial notch of the protocristid as on IRNSB M 1474 (Figure 16). On IRNSB M 1474, the inclusion of the metaconid to the talonid can be explained by the broken protocristid (effect of the wear facet 1) and strong development of the cristid obliqua. On IRNSB M 1474, the hypoconid and hypoconulid are more or less fused in disto-lingual position as on some M2 of Le Quesnoy. On the M1 of ambloctonines, the talonid cusps are more developed and separated. The talonid is also wider and more basined. IRNSB M 1474 and the M2 of P. gigantea of Le Quesnoy also share: a metaconid more reduced than on M1 and posterior to the protoconid, a prefossid well opened, a pre-, ecto- and postcingulid, and a lingual cingulid running from the base of the paraconid to the apex of metaconid. The similarities of size, wear pattern and morphology with Le Quesnoy material support the identification of IRNSB M 1474 as an M2 of Palaeonictis gigantea. Dormaalodon woutersi is here recognized as a junior synonym of Palaeonictis gigantea. DiscussionDe Blainville (1842) has described the first Ambloctoninae, Palaeonictis gigantea from a mandible MNHN ARP52 found near Paris in the "Argiles à lignites du Soissonnais". Even though some other specimens have been found during the XIXth century in the Paris Basin (Rat 1965), the European ambloctonines are very rare and poorly studied. The most recently described specimens of Palaeonictis gigantea (one P4 and one DP4) have been found in the Belgian localities of Dormaal and Hoegaarden (Smith and Hooker 1996; Smith and Smith 2001). Our study of the material previously described from "Argiles à lignites' (MNHN ARP 52, MNHN ARP 53, MNHN ARP 54), Meudon (MNHN ARP 6) and Pourcy (MNHN L-Py-60) shows that these specimens are similar to the fossils found in Le Quesnoy. Contrary to the Oxyaena material, there are no distinctive characters which justify a specific separation of specimens from localities younger than Le Quesnoy (such as Meudon, Muirancourt and Pourcy). The specimens from these younger localities are slightly larger than those of Le Quesnoy. They also differ from the material of Le Quesnoy by a few features (e.g., metaconid more distal on M1, more pronounced labial cingula on premolars and molars) that correspond to trends observed in the North American ambloctonines. Three genera of Ambloctoninae are known in North America: Dipsalodon, Palaeonictis and Ambloctonus. Those genera form a lineage characterized notably by the reduction of the M2 and of its talonid, by lower molar trigonid more secant with a reduction of the metaconid and by a molarization of the P4. Some of these features are probably linked to an omnivorous or durophagous diet. The M2 found in Le Quesnoy shows a reduced talonid and metaconid much more similar to Palaeonictis than to Ambloctonus and Dipsalodon. Four species of Palaeonictis are known: P. gigantea (MP7) from Europe; Palaeonictis peloria Rose, 1981 (Cf3), Palaeonictis occidentalis Osborn and Wortman, 1892 (Wa1-Wa4) and the recently described Palaeonictis wingi Chester et al., 2010 (Wa0) from North America. P. peloria is the oldest and biggest, and P. wingi is the smallest (Rose 1981; Chester et al. 2010). The P4 found in Le Quesnoy is morphologically close to P. gigantea known from Dormaal (IRSNB M1355) and the Paris Basin (MNHN ARP 52). It is slightly longer (almost 10%; Table 6 and Table 7) than those of Dormaal and Paris Basin. The P4 MNHN QNY2-2560 has a talonid slightly longer relative to the protoconid than seen in IRSNB M1355 and P4 of the type of P. gigantea. The P4 MNHN QNY2-2556 is more similar to the last two specimens. These slight differences do not justify the erection of a new species because the talonid of the P4 seems to be variable and because we do not know the variation of size of Palaeonictis from Dormaal and the "Argiles à lignites du Soissonnais". The talonid of the M1 from Le Quesnoy is very similar to the type specimen studied by de Blainville (1842). The larger size of Le Quesnoy fossils is within the size variation of the Ambloctoninae. In P. occidentalis, the difference in size can reach 16% for the M1 (calculated after Rose 1981, table 34). Rose (1981) and Smith and Smith (2001) proposed to root P. gigantea in the Paleocene P. peloria (Cf3), the older and most plesiomorphic species known in North America for this genus. Chester et al. (2010) proposed that P. peloria gave rise to P. gigantea and P. wingi. • The P4 is smaller compared to M1 than in P. occidentalis and is also less molarized. The P4 of P. gigantea is morphologically similar to P. peloria. • In P. gigantea the ratio of M1/M2 is close to 1.36. This is intermediate between P. peloria (1.14) and P. occidentalis (1.34-1.44). This ratio is close to those calculated for P. wingi (1.26 for left M1-M2 and 1.34 for right M1-M2). The reduction of the size of the M2 is less important in P. gigantea and P. wingi than in P occidentalis. • The entoconid of the M1 is less developed than in P. peloria, but more developed than in P. occidentalis. The entoconid of M1 tends to reduce in Ambloctoninae. P. gigantea and P. wingi differ from P. occidentalis by less developed labial cingulids on M1. This feature is primitive. • P. gigantea and P. peloria are similar in the lesser reduction of the talonid of the M2 and metaconid of M1 and M2, and in the narrower cheek teeth compared to P. occidentalis. P. wingi differs from P. gigantea and P. peloria by a more reduced metaconid on M2 (Chester et al. 2010). In P. gigantea the talonid of the M2 is less reduced, and the talonid of the M1 is narrower and shorter than in P. occidentalis (Wa1-Wa4). The talonid reduction in M2 and enlargement in M1 are trends in the evolution of the Ambloctoninae. In these regards, P. gigantea is slightly more derived than P. peloria. However, P. gigantea remains less advanced than P. occidentalis for all the following characters. • The parastyle of P4 and M1 is more reduced than in P. peloria but less than in P. occidentalis. • In correlation to the wider talonid of P4 and M1, the protocone of P4 and M1 are larger and more elongated mesio-distally than in P. peloria. The entoconid on P4 is less important than on P. occidentalis.
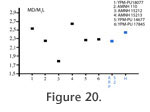
P. gigantea is, in accordance to Rose (1981) and Smith and Smith (2001), closer to P. peloria than to P. occidentalis (Figure 18). It is more derived (e.g., reduction of the M2, reduction of the talonid of the M2) and smaller than the earlier P. peloria. Its size and features (e.g., reduction of the M2) indicate a close relationship with the recently described P. wingi. The two species, in accordance to v, originated in P. peloria. The small size of the two species could have been the result of an evolutionary "Dwarfing" event that occurred during the PETM (see Chester et al. [2010] for a discussion).
Description of the Referred Postcranial Material from Le Quesnoy Forelimb. MNHN QNY2-2634; right distal humerus. This humerus is larger than the humerus of Oxyaena woutersi (QNY2-2526). Only a distal fragment is known. The medial epicondyle area is broken. In medial and lateral views, the fragment appears to be almost flat. The trochlea is slightly projected anteriorly. The elbow joint is weakly elongated transversally. The trochlea is well developed and bears a deep and concave posterior trochlear groove. The capitulum is small and almost flat. The medial lip of the trochlea is high. In anterior view, there is only a wide and transversally elongated coronoid fossa. The radial fossa is not visible (fused with the coronoid fossa?). No foramen is present on the fossa. The pit for attachment of an ulnar collateral ligament is deep and clearly visible in posterior view. The olecranon fossa is deep and transversally wide. MNHN QNY2-2639; right radius. The radius is complete. The distal epiphysis is unfused, which indicates a juvenile individual. The diaphysis is slightly twisted. The distal part is more robust than the proximal part. The radial head is oval and transversally elongated. The capitular eminence is present, elongated and not very salient proximally. The ulnar proximal facet is slightly concave. There is a well-developed bicipital tuberosity. The area for insertion of the pronator teres is well developed and is curved. The ulnar distal facet is present and slightly concave. The distal part of the radius is more triangular and massive. It is elongated mediolaterally and is moderately deep and concave – the deeper part being near the styloid process. The styloid process is low. The tubercle of the radius (origination of the abductor pollicis longus) is well developed dorsodistally. Hindlimb. MNHN QNY2-2604, MNHN QNY2-2605; two left astragali. Two complete astragali are known. The astragalus is slightly elongated anteroposteriorly and appears to be robust. The astragalar trochlea for the articulation with the distal tibia is very poorly grooved. The body is wide mediolaterally and short anteroposteriorly. The neck is slightly narrower. It is also very short anteroposteriorly. The head is anteroposteriorly wide and short. It is convex anteriorly. It is less flattened dorsoventrally than in the astragalus of Oxyaena. The astragalar foramen is weakly shifted posteriorly. In medial view, the medial part is lower than the lateral one. The medial surface is laterally extended. In dorsal view, the medial side is salient medially: this could represent a spiral facet which articulates with the medial malleolus of the tibia. On ventral view, the sustentacular facet (convex) appears to be wide and rounded. It is not extended anteroposteriorly. It is well separated from the navicular facet. Its disto-medial part is distinctly inclined ventrally. The ectal facet is strongly concave. It is wide and very short anteroposteriorly. The sustentacular and ectal facets are close. Lateral to the ectal facet, there is a deep (but transversally short) groove for the tendons of the plantaflexor muscles. The navicular facet is well extended transversely. It is rotated medio-ventrally relative to the body. In MNHN QNY2-2604, a third articular facet with the calcaneum is visible ventrally, along the navicular facet. DiscussionThe postcranial bones of Palaeonictis occidentalis are well known. Sinclair and Jepsen (1929) described an almost complete specimen (number 13001 housed at the Geological Museum of Princeton University, now in Yale). The second species of Palaeonictis (P. peloria) is only known by dental material. v concluded that Palaeonictis was a robust terrestrial ambulatory predator. The postcranial elements of Palaeonictis and Oxyaena are nearly similar (Sinclair and Jepsen 1929; Rose 1990). Our discrimination of the different postcranial elements of Oxyaenidae found in Le Quesnoy is based on size differences. P. gigantea is here compared to the oldest Oxyaena and Dipsalidictis species. The humerus is larger than that of Oxyaena woutersi but is similar in morphology. It differs by a slightly steeper trochlea. The articulation with the head of the radius is antero-posterior deep. The great capitular eminence is well developed on the radius, so the humeroradial articulation is less flexible than in Oxyaeninae (Rose 1990). The deep radial and olecranon fossae on the humerus indicate an articulation with large antero-posteriorly mobility. All these features imply that the humeroulnar joint had a strong stability. The radius is smaller than the one published by Sinclair and Jepsen (1929) (102.17 vs. 104 mm long). This is not surprising because P. gigantea is slightly smaller than P. occidentalis. The diaphysis of the radius is straighter than in Oxyanea and Dipsalidictis, probably linked to its increased weight (and robustness). However, the diaphysis in Palaeonictis is slightly twisted as in the primitive species. The head is more ovoid (so more derived) than in Dipsalidictis (rounder) and Oxyanea (Rose 2001), which limits the degree of pronation and supination (Sinclair and Jepsen 1929; Denison 1938). The radius appears to be designed more for heavy weight-bearing. The distal part is wider anteroposteriorly and flatter than in Dipsalidictis, which increases the articulation of the radius with the carpus. The facet of articulation with the ulna is deeper and more developed, indicating a strong contact between the two bones and a limitation of the movement between them. The astragalus is similar to what is known in P. occidentalis. The astragalus has a shallow and short trochlea. It is suprising that there is so little antero-posterior movement at the ankle. This morphology does not prevent entirely small rotatory movements. This conformation could be a primitive feature. The broad and dorsoventrally compressed head and the short neck may be linked to a plantigrade locomotion (Denison 1938; Ginsburg 1961; Rose 1990). Moreover the neck of the astragalus is shorter in plantigrade species than in digitigrade species. The more robust and weight-bearing morphology is not surprising because Palaeonictis is noticeably larger than Oxyaena, and implies more robust postcranial elements. As previously indicated, Sinclair and Jepsen (1929) considered Palaeonictis as a plantigrade and robust terrestrial ambulatory predator. This result agrees with that obtained for Oxyaena woutersi (see above). The ancestors of Oxyaenidae are presently poorly known (no postcranial material has been found for the Paleocene Tytthaena), but based on the data available for Dispsalidictis, they could have been arboreal. The short limbs of Oxyaenidae (e.g., Palaeonictis) might not be necessarily an adaptation to semifossorial habits as proposed by Heinrich and Rose (1995), but rather retention of primitive and arboreal features. The robust morphology of Palaeonictis and its joints indicate that P. gigantea was not cursorial. However, Palaeonictis gigantea appears – as does Oxyaena – to have a combination of arboreal (and primitive) and terrestrial features. The astragalus of P. gigantea does not show more adaptation toward a terrestrial locomotion than in O. woutersi. However, the elbow joint appears to be more restricted than in O. woutersi, which supports an ambulatory terrestrial locomotion for Palaeonictis gigantea. The calcaneum morphology of P. gigantea would possibly also reflect more limited ankle movements. According to our weight estimation, P. gigantea approaches 10 kg. The Le Quesnoy's flora indicates a wet river forest habitat surrounded by semi-deciduous or deciduous woodlands (Nel et al. 1999). It is possible that Palaeonictis gigantea and Oxyaena woutersi were able to move on the ground as easily as in the trees (thanks to primitive features). Because of the diversity of environments in Le Quesnoy, the presence of various locomotor abilities (derived terrestrial abilities and primitive arboreal abilities) was probably an advantage for the Oxyaenidae.
Palaeonictis cf. gigantea
MNHN Hou 01; right mandible with alveoli of P1, P2, M1 and M2, and very worn P3 and P4. Locality. Exact locality unknown. Found near Houdancourt, which is 2.4 km from Le Quesnoy. Argiles à lignites du Soissonnais, MP7. DescriptionMeasurements: Table 9 Dentary and lower dentition: Only a right mandible has been found. The size and number of the teeth indicate that they correspond to permanent teeth. The dental alveoli for the canine, P3, P4 and the anterior part of the M1 and M2 are preserved but are very worn. All the teeth are closely inserted. The canine alveolus is very large. It extends distally far below the P2. Except for the single-rooted P1, all the premolars are two-rooted. The size of the premolars increases from P1 to P4. The P4 is longer than the M1. The M1 is wider than the M2. Teeth form a parabolic row in the horizontal ramus, with the P3 and P4 very labially located. The canine is inserted close by the P1 and P2 and pushes them lingually.The horizontal ramus is high and robust. Its height slightly increases posteriorly. The symphysis is deep and massive, and extends between the P2 and P3. Three mental foramina are visible: below the crown of P2, below the two roots of P3 and below the anterior root of P4. The anterior mental foramen marks the beginning of a mandibular canal. The alveolar canal is present ventro-labially to the alveolus of the canine. The distal opening of the mandibular canal is seen in the ventral part of the masseteric fossa. Anterior parts of the coronoid crest and masseteric fossa are present. The coronoid crest is worn but seems to have been strong and high. The M2 extends to the coronoid crest. The masseteric fossa is deep. DiscussionNumerous characters of Palaeonictis are visible on the mandible: the parabolic shape of the tooth row in occlusal view which is typical of the genera Palaeonictis and Ambloctonus by contrast to Dipsalodon where it is more straight; the dental formula (one large canine, four premolars of which the first one is single-rooted, and two molars); the relative size of the teeth (P4 as robust as M1 and M1 longer and wider than M2); and the mandible robust and deep. The two teeth present on the mandible are too worn to be described in detail. The teeth are slightly larger than in Le Quesnoy and in de Blainville's (1842) samples. The teeth are slightly smaller than in P. occidentalis but more closely placed. They are slightly more closely located than the primitive P. peloria and Dipsalodon. The mandible shares with P. peloria, P. occidentalis and Ambloctonus the particular insertion of the P2 which is a little transverse to the axis of the tooth row. The P1 is more lingual than in P. peloria and P. occidentalis because of the proximity of the canine. This insertion gives a more parabolic aspect to the tooth row. The symphysis is shorter on the French fossil: it extends below the anterior root of P3 (as in Dipsalodon matthewi Jepsen, 1930) whereas it is below the posterior root in P. occidentalis. P. peloria has a more robust mandible with a longer symphysis than P. occidentalis. The P2 and P1 are more developed than in P. occidentalis. These features are primitive for the ambloctonines. As for P. occidentalis, a mental foramen is located under the P3.
This specimen is referred to as Palaeonictis cf. gigantea because it is the only species of Palaeonictis known in Europe and because P1 and P2 are wider than in P. occidentalis (primitive feature). The specimen is not referred formally to P. gigantea because of its larger size compared to other specimens referred to P. gigantea and because the morphology of the teeth is unknown. The Houdancourt specimen of P. cf. gigantea retains some primitive characters (e.g., P1 and P2 not reduced), but has also some derived characters (compression of the tooth row). These could be the peculiar characters of the European Ambloctoninae. Conclusion on the European AmbloctoninaeRussell et al. (1982) listed Oxyaena and Palaeonictis in the faunal list of Mutigny (MP8+9). However, our study of the carnivorous mammals of this locality refutes this occurrence. All the material appears to represent hyaenodontids. As for the Oxyaeninae, the Ambloctoninae were poorly known in the Early Eocene of Europe. They are documented by fragmentary material. The study of this new material, mostly from Le Quesnoy, allows a better understanding of the evolution of this group in Europe. Our demonstration of the synonymy of Dormaalodon with Palaeonictis implies that the Ambloctoninae were only represented by the genus Palaeonictis. By contrast to the Oxyaeninae, there is no support for an endemic evolution of the Ambloctoninae in Europe. A single dispersal of the Ambloctoninae from North America to Europe is suggested during the Paleocene-Eocene boundary, followed by an endemic evolution during the beginning of the Early Eocene, as for the Oxyaeninae. The European evolution of Palaeonictis is characterized by a slight size increase, and by morphologically similar trends to what is known in the North American lineage of P. occidentalis (e.g., paraconid more lingual, metaconid more distal, protoconid lower, P4 with less individualized entoconid). European Palaeonictis evolved convergently with North American lineages. However, by contrast to North American Ambloctoninae, which diversified and persisted until the end of the Early Eocene, the European Ambloctoninae remained restricted to one genus and rapidly disappeared. |
|

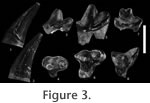
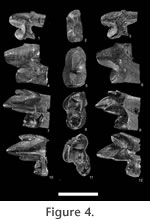
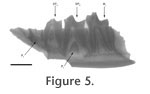

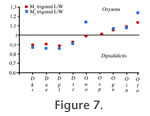
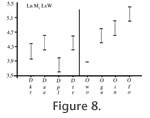
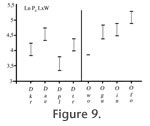 The peculiar development of the paraconid (base developed lingually and shifted mesially) of the molars of O. woutersi is shared with D. transiens and O. gulo. They also share a long postmetacrista. They share a more robust P3 and much
more developed cingula around the protocone than in other Dipsalidictis species. All these features are indicative of affinities with the genus Oxyaena. O. woutersi also shares with D. transiens a more pronounced ectoflexus on the upper molars. We think that the species of Dipsalidictis closest to O. woutersi is D. transiens. Both O. woutersi and D. transiens present notably a development toward secant lower and upper molars. O. woutersi differs from D. transiens by a more derived dentition toward carnivory (e.g., paracristid more developed and metaconid more reduced on M2), and a P3 and P4 with a talonid more robust as in Oxyaena (derived features).
The peculiar development of the paraconid (base developed lingually and shifted mesially) of the molars of O. woutersi is shared with D. transiens and O. gulo. They also share a long postmetacrista. They share a more robust P3 and much
more developed cingula around the protocone than in other Dipsalidictis species. All these features are indicative of affinities with the genus Oxyaena. O. woutersi also shares with D. transiens a more pronounced ectoflexus on the upper molars. We think that the species of Dipsalidictis closest to O. woutersi is D. transiens. Both O. woutersi and D. transiens present notably a development toward secant lower and upper molars. O. woutersi differs from D. transiens by a more derived dentition toward carnivory (e.g., paracristid more developed and metaconid more reduced on M2), and a P3 and P4 with a talonid more robust as in Oxyaena (derived features).

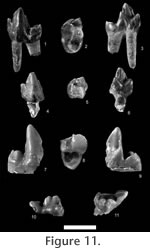


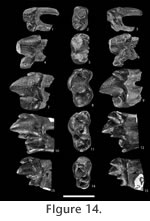
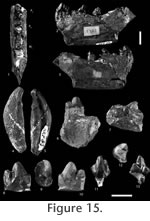
 Referred material from Pourcy.
Referred material from Pourcy.
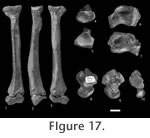
 P. gigantea and P. wingi are both characterized by a small size compared to P. peloria and P. occidentalis. However, P. gigantea differs from P. wingi by a more reduced P3 (derived feature also present in P. occidentalis; see
P. gigantea and P. wingi are both characterized by a small size compared to P. peloria and P. occidentalis. However, P. gigantea differs from P. wingi by a more reduced P3 (derived feature also present in P. occidentalis; see