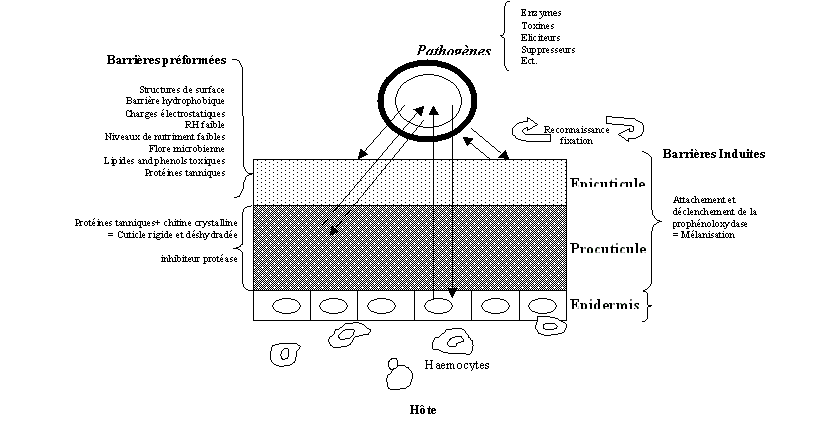
Ils sont naturellement présents dans lenvironnement (sol, air, eau) et infectent généralement leur hôte soit par ingestion, par la cuticule ou par les orifices. Le pathogène se multiplie dans lhôte en lui causant des dommages par destruction des tissus, par septicémie ou toxémie entraînant sa mort plus ou moins immédiate. Tous ces micro-organismes possèdent des formes de résistance leur permettant de persister dans lenvironnement et de perpétuer leur cycle de vie (Jourdheuil et al., 1992). Cependant, les micro-organismes ont des spectres daction assez étroits à cause de certains facteurs abiotiques qui peuvent être limitant pour leur développement optimal. Par exemple les UV (Ignoffo et Hostetter, 1977; Burges, 1981; Roberts et Campbell, 1977; Gardner et al., 1977; Fargues et al., 1988), les variations suboptimales de température (Roberts et Campbell, 1977; Doberski, 1981; Stathers et al., 1993) et de lhumidité (Fargues, 1972; Ramoska, 1984; Riba et Marcandier 1984; Khachatourians, 1987) sont détrimentales pour la plupart des micro-organismes.
1. Les bactéries
Selon Starnes et al., 1993, plus dune centaine de bactéries ont été identifiées comme ayant un potentiel dutilisation en lutte biologique. Ces bactéries entomopathogènes appartiennent surtout à trois grandes familles qui sont les Bacillaceae, Enterobacteriaceae et Pseudomonaceae (Greathead et al., 1994). À lheure actuelle, Bacillus thuringiensis Berliner et B. sphaericus sont les espèces les plus utilisées en lutte contre les ravageurs. Pour B. sphaericus, la toxine est localisée dans la paroi sporale et va être libérée par une digestion partielle de la bactérie dans le tube digestif de la larve de linsecte. La toxine pénètre dans la membrane péritrophique du tube digestif et empoisonne la larve (Burgess, 1981; Singer, 1981). Les bactéries se développent dans lhôte et le quittent quand celui-ci se désintègre. Nous développerons davantage le cas du B.t à cause de sa grande utilisation tant en agriculture quen foresterie et en milieu aquatique. Certaines souches de B.t possèdent une spore et une inclusion parasporale composée dune ou plusieurs toxines protéiques. Quatre types de toxine peuvent être isolées du B. thuringiensis, les ![]() -exotoxines (Heimpel, 1955; Heimpel, 1967; Kushner et al., 1957; Toumanoff, 1953); ß-exotoxines (Burgerjon, 1965; Heimpel, 1967; Heimpel et Angus, 1963);
-exotoxines (Heimpel, 1955; Heimpel, 1967; Kushner et al., 1957; Toumanoff, 1953); ß-exotoxines (Burgerjon, 1965; Heimpel, 1967; Heimpel et Angus, 1963); ![]() -endotoxines et ÿ-exotoxines (Heimpel, 1967; Heimpel et Angus, 1963). Dans le mésenteron de linsecte, en présence dun pH stomacal basique (8,9), le cristal va se désintégrer pour libérer une endotoxine, un polypeptide toxique qui va provoquer une rapide baisse de lATP au niveau des cellules stomacales, un gonflement de cellules epithéliales, une paralysie du tube digestif et un débalancement ionique dans lhémolymphe. Linsecte meurt par inanition (Fast, 1981; Cloutier et Cloutier, 1992; Greathead et al., 1992; Miller et al., 1983) ou par une septicémie provoquée par la multiplication de la bactérie dans lhemolymphe et les tissus.
-endotoxines et ÿ-exotoxines (Heimpel, 1967; Heimpel et Angus, 1963). Dans le mésenteron de linsecte, en présence dun pH stomacal basique (8,9), le cristal va se désintégrer pour libérer une endotoxine, un polypeptide toxique qui va provoquer une rapide baisse de lATP au niveau des cellules stomacales, un gonflement de cellules epithéliales, une paralysie du tube digestif et un débalancement ionique dans lhémolymphe. Linsecte meurt par inanition (Fast, 1981; Cloutier et Cloutier, 1992; Greathead et al., 1992; Miller et al., 1983) ou par une septicémie provoquée par la multiplication de la bactérie dans lhemolymphe et les tissus.
Le B.t est efficace contre certaines espéces de coléoptères, lépidoptères et diptères (Morris, 1983; Ahmed et al., 1994). Cependant il ne serait pas efficace contre les acridiens en raison de lacidité intestinale (Greathead et al., 1994). Le B.t ne doit pas oblitérer lutilisation dautres bactéries qui se sont révélées efficaces comme le genre Xenorhabdus (Enterobacteriaceae), très connu pour sa virulence contre les acridiens. Elles forment le plus souvent des symbiotes avec les nématodes qui agissent comme vecteur. Libérées dans lhomocoele, elles tuent rapidement leur hôte (Greathead et al., 1994). B. popillae est efficace contre les Scarabaeidae et a été utilisé avec succès contre le scarabée japonais Popillia japonica (Hutton et Burbutis, 1974; Miller et al., 1983; Krieg, 1987). Dans le cas de B. popillae, les spores ingérées germent dans le tube digestif et traversent la membrane epithéliale, linsecte meurt par septicémie (Burgers, 1982). Pseudomonas aeruginosa et Serratia marcescens (Saik et al., 1990) sont des agents potentiels de lutte. Lutilisation répétée des bactéries peut toutefois, comme les pesticides chimiques, entraîner une résistance chez certaines espèces (Dunphy et Tibelius, 1992).
2. Les Virus
Les virus entomopathogènes se divisent généralement en deux grands groupes distincts, dune part, ceux possédant des corps dinclusion paracristallin et ceux sans corps dinclusion. On les regroupe en sept familles. Ce sont, les Baculoviridae, Reoviridae, Poxviridae (à corps dinclusion); les Iridoviridae, Parvoviridae, Picornoviridae et les Rhabdoviridae (sans corps dinclusion) (Faulkner et Boucias, 1985; Arata et al., 1978). Ces familles renferment la plupart des 650 espèces de virus entomopathogènes connues (Khachatourians, 1986). Ce sont les Baculoviridae, les Reoviridae (Miller et al., 1983) et les virus entomopox (poxviridae) qui sont les plus utilisés en lutte biologique, car ils sont bénins pour les vertébrés, les corps dinclusion ne pouvant se développer que chez les insectes (Paynes, 1982). Les baculovirus ont depuis longtemps présenté un intérêt principalement pour leur spécificité. Ils nont en effet été observé que chez les invertébrés et en particulier chez les insectes (Devauchelle, 1993). Ce sont des virus en bâtonnet dont le génome est constitué dune molécule dADN bicaténaire de haut poids moléculaire (70-85 106 daltons). Quant aux entomopox, ils sont parasites de plusieurs acridiens ravageurs de cultures (Purrini et al., 1988) et sont comme les baculovirus, inoffensifs pour les vertébrés, doù leur grand intérêt en lutte biologique (Street et Mc. Guire, 1990). Les virus sont des parasites obligatoires et ne peuvent se reproduire que dans les cellules animales ou végétales. Au cours du processus dinfection dans le noyau des cellules, ces virus forment des corps dinclusion appelés polyèdres qui sont constitués de nombreuses particules virales dans une matrice protéinique composée principalement dun simple polypeptide, la polyedrine. Les polyèdres ingérés vont être dégradés par les protéases du tube digestif de linsecte et les virions libérés traversent les cellules intestinales pour se multiplier dans les hemocytes et dans les tissus adipeux. Il est rapporté par Meynadier et al., (1993) que dans certains cas, les virus liquéfient les corps gras entraînant une turgescence de linsecte suivi de sa mort. Les polyèdres formés dans le noyau provoquent la polyedrose nucléaire (NPV) et affectent principalement les lépidoptères et hymenoptères tandis que les baculovirus à corps dinclusion granulaire (GV) causent la granulose et affectent surtout les lépidoptères.
Au niveau des rickettsies, les publications sont plus rares. Cependant Vago et Martoja (1963) ont mentionné quune rickettsie, Rickettsiella grylli existait chez les grillons et était capable de contrôler leur population.
Les caractéristiques principales des bioinsecticides viraux sont la spécificité, la haute virulence, la rapidité daction et le niveau raisonnable de persistance dans lenvironnement (Dent, 1991). La rémanence des virus est cependant affectée par les radiations UV. Par exemple les NPV et GV sont inactivés après quelques heures dexposition au rayonnement solaire (Franz, 1971).
3. Les Protozoaires (Grégarines, Microsporidies et Amibes)
Les protozoaires appartiennent à sept phyla, dont quatre, les Ciliophora, Sarcomastigophora, Apicomplexa et Microspora sont pathogènes des insectes (Dent, 1991). Les familles les plus utilisées en lutte biologique sont les Amoebidae et les Nosematidae (Greathead et al., 1994). Par exemple Bralamoeba locustae (Amoeba appartenant au phylum des Sarcomastigophora) est très efficace pour le contrôle des locustes (Canning, 1982). Parmi les néogrégarines, ce sont les microsporidies qui offrent le plus de potentiel en lutte microbiologique en tant quorganismes unicellulaires eucaryotes (Canning, 1982). Ce sont des parasites intra-cellulaires obligatoires qui forment des spores caractéristiques. Chez les microsporidies du genre Nosema, linfection se réalise par ingestion des spores, celles-ci germent dans le tube digestif et traversent les tissus éphitéliaux (Maddox, 1987). Dans le cytoplasme, lagent infectieux se multiplie en produisant dautres spores qui vont envahir linsecte et peuvent par transmission verticale infecter la génération suivante. Les protozoaires provoquent des maladies chroniques à évolution lente ou des enzooties, qui affaiblissent et affectent la croissance ou la fécondité de leur hôte plutôt que dentraîner une mort rapide (Poinar et al., 1985; Cloutier et Cloutier, 1992). Cependant lhôte infecté devient souvent plus sensible à dautres infections dorigines virales, bactériennes ou mycoses (Khachatourians, 1986). Lespèce Nosema locustae a été utilisée avec succès dans la lutte contre les acridiens aux USA. Une mortalité de plus de 95% a été obtenue avec une infection précoce des larves (Greathead 1992b; Kevan, 1992). Des mortalités ont été obtenues avec Vairimorpha necatrix en infectant des larves de lépidoptères particulièrement des noctuidae. Dans les régions sahéliennes dAfrique et les régions arides dAsie, N. locustae Canning est très prometteur dans la lutte anti-acridienne en raison des possibilités de production et de dissémination par ultra bas volume et de son innocuité vis-à-vis des vertébrés. Les microsporidies Nosema pyrausta et Vairimorpha necatrix sont potentiellement exploitables contre les ravageurs des cultures localement importantes (Andreadis, 1987). La revue de littérature est ambiguë sur lutilisation des microsporidies. Nous avons deux pôles divergeants; dune part, ceux qui ne considèrent pas les microsporidies comme des agents de lutte biologique efficaces car nentraînant pas la mort de lhôte mais plutôt des dommages tels les effets débilitants ou affaiblissants et dautre part ceux qui soutiennent que ce sont des agents de lutte efficace. Il est donc nécessaire de lever cette équivoque. En tout état de cause, en tant que parasite intracellulaire obligatoire, les microsporidies tout comme les virus sont très difficiles à multiplier de façon massive pour la production de biopesticides (Khachatourians, 1986; Poinar et al., 1985). Cependant des formulations commerciales existent.
Parmi les Amoebidae, Melameba locustae King et Taylor est connu comme agent pathogène pouvant conduire à la mortalité des criquets et des grillons (Kevan, 1992). Il infecte principalement les tubes de Malpighi des criquets (Kevan, 1992) et de certains tétrigidés et gryllidés (Taylor et King, 1937; Lea, 1958; Prinsloo, 1960, 1961a; Dempster, 1963; McLaughlin, 1971; Henry et Oma, 1981; Braun et al., 1988; Streett et McGuire, 1990). Melameba locustae semble celui de tous les protozoaires amiboïdes entomophiles qui présente le plus dintérêt en lutte microbiologique (McLaughlin, 1971). Quant aux grégarines, on les retrouve particulièrement chez presque tous les groupes dorthopteroïdes (Steinhaus, 1949; 1963; Gunter et Herter, 1974). Ce sont les protozoaires intestinaux les plus rependus chez les insectes et particulièrement chez les criquets (Geathead, 1966a; Theodoridès et al., 1958; Steinhaus, 1949). Les dommages quils infligent à lépithélium intestinal permettent à dautres bactéries entomopathogènes denvahir lhôte et de provoquer des septicémies (Dempster, 1963). Harry, (1970) a montré que leugrégarine Gregarina garnhami, empêchait les imagos de criquets pèlerins Schistocerca gregaria de prendre du poids. Pour McLaughlin, (1971), ces protozoaires sont potentiellement utilisables en lutte biologique bien que plusieurs auteurs ne soient pas de cet avis.
4. Les nématodes
Il existe plusieurs espèces de nématodes parasites dinsectes. Pour la plupart delles, linfection se fait à partir dufs déposés sur les feuilles des plantes. Les ufs éclosent et les larves regagnent lhomocèle et au quatrième stade quittent lhôte par perforation des tissus intersegmentaires. Il sen suit la mort de linsecte. Certaines espèces de Steinermatidae et Heterorhabditidae vivent en symbiose avec des bactéries du genre Xenorhabdus. Les larves pénètrent lhôte par les orifices naturels et même par la cuticule ou elles libèrent les bactéries qui tuent rapidement lhôte. Quoique de bons agents en lutte biologique, lutilisation des nématodes en zone sèche est limitée par les facteurs abiotiques particulièrement les UV qui sont détrimentales pour tous micro-organismes (Ignoffo et Hostetter, 1977; Burges, 1981; Roberts et Campbell, 1977; Gardner et al., 1977) et peuvent entraver le processus dinfection de lhôte (Greathead et al.1994).
5. Microchampignons
Parmi les micro-organismes utilisés en lutte biologique, plus de 700 espèces de microchampignons sont entomopathogènes (Starnes et al.; 1993) et jouent un rôle important dans la régulation naturelle des populations dinsectes (Wraight et Roberts, 1987; Ferron, 1978). Ils appartiennent au sous-taxon des Mastigiomycotina, Zygomycotina, Ascomycotina et Deuteuromycotina. Le plus grand nombre de pathogènes se trouvent dans la classe des Zygomycètes, mais les plus utilisées en lutte biologique proviennent des Deuteromycètes (Fungi imperfecti). Les espèces des genres Beauveria, Metharizium, Verticillium, Erynia, Hirsutella, Entomophtora et Entomophaga sont les plus utilisées en lutte biologique (Wraight et Roberts, 1987; Goettel, 1992). Ils ont un intérêt agronomique considérable dans la lutte biologique contre les ravageurs de cultures et sont donc lobjet détudes de plus en plus poussées. La pathogénécité de l'inoculum sporal et la spécificité de l'hôte sont deux paramètres important dans le choix de l'isolat fongique. Les microchampignons entomopathogènes sont des agents de lutte très intéressant du fait de leur aptitude à infecter l'hôte par ingestion ou par simple contact rendant tous les stades, uf, larve, adulte sensibles ainsi que les succeurs-piqueurs (Carruthers and Soper, 1987). Ils peuvent être produits en masse à moindre coût et peuvent être appliqués avec les méthodes conventionnelles. Les principaux facteurs limitant l'utilisation en champ des microchampignons sont abiotiques et vont entraîner la perte d'efficacité de l'inoculum fongique sur le couvert végétal. Les effets de certains facteurs sur la viabilité des conidies ont été très étudiés comme la température (Roberts et Campbell, 1977; Doberski, 1981; Stathers et al., 1993), l'effet du rayonnement solaire sur la rémanence ou l'inactivation de l'inoculum infectieux (Ignoffo et Hostetter, 1977; Burges, 1981; Roberts et Campbell, 1977; Gardner et al., 1977), l'effet de l'humidité (Fargues, 1972; Hall et Papierok, 1982; Ramoska, 1984; Riba et Marcandier 1984; Khachatourians, 1987).
Le champignon entomopathogène Beauveria bassiana
Historique et taxonomie
Parmi les micro-organismes entomopathogènes ayant un potentiel dagent de lutte biologique contre les insectes nuisibles, plus de 500 espèces de champignons sont susceptibles d'infecter des insectes (Starnes et al., 1993). Metarhizium anisopliae était le premier pathogène utilisé délibérément pour le contrôle dinsecte ravageur par le Russe Eli Metchnikoff (1880) (le père de la lutte microbiologique) dans les années 1880. B. bassiana (Hyphomycète) est un microchampignon pathogène pour de nombreux insectes. Sa pathogénicité a été démontrée pour la première fois par Agostino Bassi de Lodi (1835), le précurseur des études des maladies infectieuses, en démontrant pour la première fois quun micro-organisme pouvait être responsable de maladie infectieuse chez lanimal. Beauveria sp. a été décrit par la suite par Jean Beauverie en 1911 sous le nom de Botrytis bassiana. Le genre a été établi par Vuillemin en 1912 et fut classé dans lordre des Hyphomycètes.
Il est important de mentionner que la classification des hyphomycètes a été sujette à plusieurs révisions. A lheure actuelle, on regroupe sous le nom de Hyphomycetes plus de 10 000 espèces appartenant à plus de 1 800 genres (Subramanian, 1983). La caractéristique principale des hyphomycètes est quils nont pas de reproduction sexuée, ce qui contraignait les taxinomistes à baser leur identification sur des caractéristiques morphoontogéniques.
Il existe deux systèmes de classification des hyphomycètes. Le premier préconisé par Saccardo (1886) est basé sur la morphologie et la pigmentation des conidies et conidiophores. Le second a été proposé par Hughes (1953) et sappuie sur le développement et la morphologie, considérant la pigmentation et la septation des conidies comme des caractères secondaires. Pour Talbot (1971), les deux systèmes de classification ne seraient pas rigoureux dans la mesure ou les caractéristiques peuvent varier et peuvent surtout changer avec les conditions de croissance et de maturité de ces champignons. Le genre Tolypocladium a été confronté très récemment au genre Beauveria (Von Arx, 1986) sur la base des similarités morphologiques. Selon lui, les variations morphologiques au niveau des phialides et des conidies ne sont pas suffisantes pour séparer les deux genres. Il a donc reclassifié toutes les espèces de Tolypocladium comme des synonymes de Beauveria. Todorova et al., (1997) à partir de tests biochimiques sur les profils dutilisation de carbohydrates (API 50 CH) ont levé léquivoque en démontrant que Beauveria et Tolypocladium sont deux genres distincts.
 |
Plusieurs corrections taxinomiques ont été apportées. Par exemple Petch (1926) et MacLeod (1954) ont distingué les espèces B. bassiana (Balsamo) Vuillemin et B. tenella (Delacroix) MacLeod, considérant notamment B. densa (Link: Fries) Picard et B. Brongniartii (Saccardo) comme des souches de B. tenella. De Hoog (1972) a proposé que les espèces B. alba (Limber) Saccas et B. bassiana sont deux espèces distinctes, considérant B. brongniartii comme identique à B. tenella et B. densa comme une souche synonyme de B. bassiana. La classification la plus complète à lheure actuelle est celle de Mugnai et al. (1989) issue détudes morphologiques et de tests enzymatiques décrivant six espèces de Beauveria. (B. alba, B. amorpha (Von Höhnel) Samson & Evans, B. bassiana, B. brongniartii, B. velata Samson & Evans, B. vermiconia (Hoog & Rao)). Une autre espèce B. caledonica (Bissett & Widden) a été décrite par Bissett & Widden (1988).
Morphologie
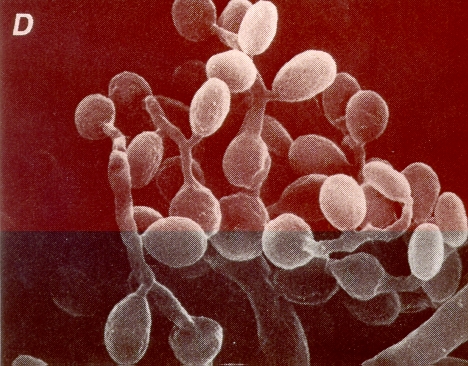
Les espèces de Beauveria produisent les colonies cotonneuses blanches à jaunâtre. Les conidies ou spores (fig. 1) sont soutenues par de long filament en zigzag qui sont des hyphes transparents et septaux (fig. 2) avec un diamètre de 2.5 à 25 µm. Les conidies sont produites sur des épis courts, donnant aux cellules conidiogènes un aspect épineux. En présence dair le champignon produit des conidiospores de forme sphérique (1-4 µm de diamètre) ou ovales (1.55-5.5 x 1-3 µm denvergure) mais en milieu anaérobie, il produit des blastospores de forme ovale (2-3 µm de diamètre et 7 µm de longueur). Les blastospores sont aussi infectieux que les conidies (Weiser, 1972, Lipa, 1975).
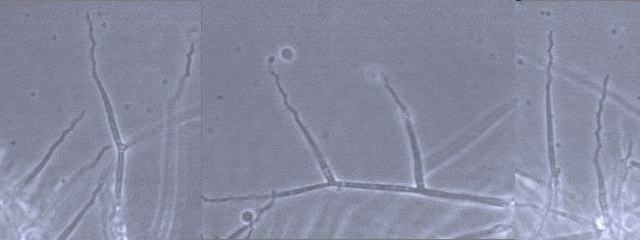 |
Le mode d'infection de B. bassiana se divise en quatre étapes distinctes qui sont l'adhésion, la germination, la différentiation; la pénétration. Ladhésion est caractérisée par un mécanisme de reconnaissance et de compatibilité des conidies avec les cellules tégumentaires de l'insecte (Vey et al., 1982) (fig. 3). Cette phase se scinde en deux étapes distinctes, la première passive ou l'attachement à la cuticule est réalisée grâce à des forces hydrophobiques et électrostatiques (Fargues 1984; Butt, 1990, Boucias et al.,1988) et la seconde active caractérisée par la production dun mucilage qui va engendrer une modification épicuticulaire (Wraight et al.; 1990) aboutissant à la germination (fig. 3). Après la phase d'adhésion, la germination va être dépendante des conditions environnantes et aussi de la physiologie de l'hôte; (composition biochimique de la cuticule de l'hôte) qui peut favoriser ou inhiber la germination (St-Leger et al., 1989; Butt et al., 1995, Smith et Grula, 1982, Butt, 1990; Butt et Becket, 1994). L'avant dernière phase est la différentiation caractérisée par la production dappressorium, structures terminales qui vont servir de point dencrage, de
ramollissement de la cuticule et favoriser la pénétration. La production des appressoria est très dépendante de la valeur nutritive de la cuticule de lhôte (St-Leger et al.; 1989; Magelhaes et al.; 1990). Une cuticule nutritive va stimuler la croissance myceliale plutôt que la pénétration (St-Leger et al.; 1989). La dernière phase est la pénétration de l'hôte qui se fait par la combinaison de pression mécanique (Pekrul et Grula, 1979) et enzymatique (Butt, 1990; Charnley, 1989; St-Leger 1993, 1995) telles que les lipases, les protéases et les chitinases (St-Leger, 1995; Segers et al., 1995; Kucera et Samsinakova 1968, Leopold et Samsinakova, 1970), la plus importante dans la pénétration étant les protéases. (St-Leger et al.; 1988; 1989; Butt et al.; 1990; Bidochka et Khachatourians; 1988). Certaines souches produisent des toxines non enzymatiques telles que la beauvericine, les beauverolides, les bassianolides, les isarolides qui accentuent et accélèrent le processus d'infection (Roberts 1981; Hajeck et St-Leger, 1994). La colonisation de l'hôte se fait lorsque le champignon parvient à surmonter les mécanismes immunitaires de défense de l'insecte (Boman et Steiner, 1981; Soderhall, 1981) et envahit l'hemolymphe (Ferron et al., 1993). À la mort de linsecte, le champignon produit un antibiotique Oosporin qui va lui permettre de surmonter la compétition des bactéries du tube intestinal de linsecte. La phase saprophyte va être caractérisée par la mummification du cadavre transformé en sclérote. Les hyphes traversent le tégument préférentiellement au niveau intersegmentaire puis le recouvre dun feutrage mycellien blanc cotonneux (fig. 4) qui va amorcer la formation des conidiospores (Weiser, 1972). On utilise le terme muscardine pour caractériser le feutrage mycellien blanc cotonneux en référence aux bonbons muscardines produit en France.
Conclusion - Avantage de B. bassiana par rapport aux autres agents de lutte microbiologique Le large spectre d'action et la virulence de B. bassiana ont permis de l'utiliser avec succès en milieu agricole en Ex-URSS et en Europe de l'Est (Lipa, 1985). Les premiers résultats positifs dans la répression des ravageurs avec B. bassiana en Amérique ont été obtenus aux USA (Roberts et al.; 1981; Campbell et al., 1985). La pathogénécité de l'inoculum sporal et la specificité de l'hôte sont deux paramètres important dans le choix de l'isolat fongique. Le microchampignon entomopathogène B. bassiana est un agent de lutte très intéressant du fait qu'il peut infecter l'hôte par ingestion ou par simple contact contrairement aux autres agents de lutte micro-biologiques. Ce mode daction particulier rend tous les stades (oeuf, larve, adulte) sensibles. B. bassiana peut être produit en masse à moindre coût et peut être appliqué avec les méthodes conventionnelles. Il n'est pas dangereux pour les vertébrés et certains isolats comme ceux utilisés dans cette thèse sont très virulents et spécifiques aux ravageurs visés, ce qui est un critère intéressant pour lentomofaune non cible. De plus, les conidies ont une assez longue persistance dans l'environnement pouvant permettre naturellement des enzooties ou épizooties.
VertigO no 2, vol 2