La lutte biologique utilise des organismes vivants pour diminuer les niveaux de population d'autres organismes, généralement nuisibles. Les ennemis naturels les plus souvent utilisés en lutte biologique comprennent les microorganismes, les nématodes, les prédateurs et les parasitoïdes. Ces derniers sont responsables de nombreux succès en lutte biologique et ils occupent dans les écosystèmes naturels une place importante. On considère que la mortalité causée par les parasitoïdes est plus importante en nature que celle attribuée aux prédateurs et aux microorganismes combinés. On retrouve des parasitoïdes dans six ordres d'insectes mais ce sont surtout les parasitoïdes hyménoptères, diptères et coléoptères qui ont été étudiés en lutte biologique. Malgré des qualités indéniables en lutte biologique (grande spécificité, innocuité pour l'homme, possibilités d'élevage de masse), l'utilisation de parasitoïdes en lutte biologique n'est pas sans problème. Les risques associés à la pollution génétique des populations naturelles et aux conditions d'élevages de masse doivent être pris en ligne de compte dans l'évaluation des coûts et bénéfices de la lutte biologique.
Le milieu agricole et le milieu forestier sont l'objet depuis plusieurs années de critiques concernant leur impact environnemental. Les citoyens s'inquiètent de la dégradation des milieux naturels ainsi que de l'innocuité des aliments qu'ils consomment. Cette perception négative a été exacerbée en Europe suite aux crises successives qui ont frappé le milieu agricole (contamination de poulets à la dioxine, crise de la vache folle, épidémie de fièvre aphteuse...). Face à ces critiques et afin de rendre moins polluante et moins risquée l'agriculture moderne, un certain nombre de solutions, dont la lutte biologique, sont proposées.
Il n'est pas inutile de définir brièvement ce que nous entendons ici par lutte biologique. Il existe de nombreuses définitions de la lutte biologique mais nous nous en tiendrons à une définition générale telle que celle proposée par Van Drische et Bellows (1996): «Biological control is a population-level process in which one species' population lowers the numbers of another species by mechanisms such as predation, parasitism, pathogenicity, or competition» (La lutte biologique est un processus agissant au niveau des populations et par lequel la densité de population d'une espèce est abaissée par l'effet d'une autre espèce qui agit par prédation, parasitisme, pathogénécité ou compétition). La lutte biologique est donc l'utilisation d'organismes vivants dans le but de diminuer la densité de population d'un autre organisme vivant, généralement un ravageur. Ce ne sont donc pas toutes les méthodes de lutte non-chimiques qui peuvent être considérées comme de la lutte biologique. Ainsi l'utilisation de produits chimiques extraits de plantes (tels des insecticides d'origine végétale) ou de microorganismes (tels les formulations de Bt ne contenant que l'exotoxine du Bt et non plus de bactéries vivantes) n'est pas de la lutte biologique. La sélection, traditionnelle ou par introduction de gènes, de plantes résistantes aux insectes ou aux maladies, la lutte culturale ou l'utilisation de produits sémiochimiques, tels les phéromones, ne sont également pas des méthodes de lutte biologique. Par contre l'utilisation de produits chimiques qui attirent ou retiennent des ennemis naturels ou la sélection de plantes qui favorisent l'efficacité des ennemis naturels contribuent à la lutte biologique.
Pourquoi s'intéresser de près à ce qui peut et ce qui ne doit pas être considéré comme de la lutte biologique? En fait, surtout pour une question de crédibilité. La montée de l'intérêt du public pour des aliments produits dans un contexte agricole sain et non-polluant pousse l'industrie agro-alimentaire à satisfaire ce besoin. Cette réponse est tout à fait souhaitable en autant que la réponse de l'industrie corresponde bien aux attentes du public. Or la lutte biologique telle que définie plus haut n'est pas simple à mettre en place et demande un effort de recherche et de vulgarisation important. La tentation peut être forte de qualifier de lutte biologique des méthodes plus faciles à mettre en place, comme par exemple le passage vers des insecticides chimiques d'origine végétale, mais qui n'apportent pas de garanties quand à leur innocuité pour l'environnement ou le public.
L'objectif de la lutte biologique est donc à terme de remplacer, en totalité ou en partie, les pesticides chimiques utilisés en agriculture et en forestrie. La lutte biologique n'est donc pas l'agriculture biologique ou organique qui demande une production complètement exempte d'intrants chimiques (engrais, pesticides, antibiotiques ...). Cependant la lutte biologique peut devenir une composante importante de systèmes de production intégrée ou biologique.
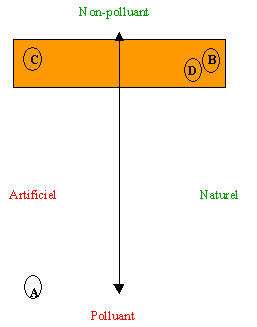 |
Figure 1- Diagramme illustrant les gradients existants en production agricole et forestière. Selon qu'une production est plus ou moins naturelle et polluante, on peut la situer sur ce plan. A- production à la fois artificielle et polluante, B et D- productions naturelles et non-polluantes, C- production non-polluante mais artificielle
L'utilisation de méthodes de lutte biologique n'est donc pas une assurance de production agricole naturelle mais plutôt d'une production moins polluante. En effet on peut placer les divers types de production agricole et forestière sur un plan séparant la notion de production polluante de celle de production naturelle (Figure 1). Sur ce graphique, la production intensive de porc en porcherie sur clayage peut être placée au point A puisqu'elle est à la fois polluante et artificielle. A l'opposé, la production de bleuet en bleuetière sauvage sans intrants est naturelle et non-polluante (point B, Figure 1). Une production utilisant la lutte biologique progressera vers la zone ombrée au sommet du graphique. La mise en place d'un programme de lutte biologique dans une production de laitue en hydroponique ne rendra pas cette culture moins artificielle mais elle sera moins polluante (point C, Figure 1). Dans le cas d'une repousse naturelle de forêt protégée contre les insectes ravageurs par des bactéries ou des parasitoïdes, on obtiendra une culture à la fois non-polluante et naturelle (point D, Figure 1). Notons au passage qu'une ferme en agriculture organique se trouvera au sommet du graphique mais pas nécessairement vers la droite. En effet la production de la plupart des fruits et légumes ainsi que la production animale peut être non-polluante mais n'est certainement pas une situation naturelle!
Terminons cette brève mise au point sur la lutte biologique en soulignant que, selon les modalités d'utilisation, on reconnaît plusieurs types de lutte biologique. On qualifie de lutte biologique classique, ou par introduction, la technique qui consiste à introduire une nouvelle espèce dans un environnement afin de contrôler les populations d'un ravageur (Pedigo 1996). La plupart du temps, cette approche vise à limiter les populations d'un ravageur exotique en introduisant un ennemi naturel provenant de la zone de distribution originale de ce ravageur mais on peut aussi tenter de transférer un ennemi naturel exotique sur un ravageur indigène. La lutte biologique inondative est une technique augmentative c'est-à-dire qu'on augmente les populations d'ennemis naturels. Dans ce cas, les quantités relâchées dans le milieu sont importantes et l'objectif est de détruire immédiatement un ravageur sans que l'établissement et la reproduction de l'ennemi naturel soient visés. L'utilisation du Bt (bactérie pathogène aux insectes) et des trichogrammes (guêpes parasitoïdes) entrent dans cette approche. Dans une autre approche augmentative, appelée lutte biologique inoculative, l'ennemi naturel est relâché en petite quantité et doit s'établir, se multiplier et coloniser une zone donnée. Cependant cet établissement n'est généralement pas permanent et des introductions doivent être faites une ou plusieurs fois par saison. La manipulation environnementale permet quant à elle de maximiser les effets bénéfiques des espèces indigènes d'ennemis naturels. L'utilisation de couverts végétaux modifiés ou de plantes nectarifères sont des exemples de cette approche.
Les organismes utilisés en lutte biologique
Pratiquement tous les organismes vivants peuvent être considérés comme des ennemis naturels selon l'angle avec lequel on examine leur écologie. Cependant dans un contexte de lutte biologique en agriculture et en foresterie, et surtout en ce qui concerne la lutte biologique contre les insectes ravageurs, quatre groupes d'organismes sont surtout utilisés. Ce sont les microorganismes, les nématodes, les prédateurs et les parasitoïdes. Nous examinerons brièvement les trois premiers groupes pour ensuite nous concentrer sur les parasitoïdes.
Les microorganismes regroupent les bactéries, virus, champignons et protozoaires pathogènes aux insectes. Les bactéries entomopathogènes se retrouvent chez les bactéries asporogènes telles les Pseudomonadaceae (Pseudomonas) et les Enterobacteriaceae (Aerobacter, Cloaca, Serratia) et les bactéries sporogènes tels les Bacillaceae (Bacillus, Clostridium). Environ une centaine d'espèces sont spécifiquement entomopathogènes (Cloutier & Cloutier 1992). Le premier groupe, les bactéries asporogènes, est présent dans le sol et souvent dans le système digestif des insectes (Van Driesche & Bellows 1996). Ce sont souvent des opportunistes qui envahissent un insecte suite à une blessure, un stress ou en conjonction avec un autre pathogène. Leur manque de spécificité et surtout leur relative pathogénicité envers les mammifères les ont évidemment tenu à l'écart des programmes de lutte biologique. Les bactéries sporogènes s'attaquant aux insectes sont plus spécifiques et spécialisées. Ce sont surtout des espèces de Bacillus telles B. thuringiensis, B. popillae et B. sphaericus qui ont été étudiées. La plupart des souches de B. thuringiensis sont virulentes contre les Lépidoptères mais quelques souches toxiques aux Diptères (B. t. israelensis) et aux Coléoptères (B. t. tenebrionis) existent. B. popillae est une bactérie pathogène aux larves de Scarabidae alors que B. sphaericus n'est pathogène que pour quelques genres de Culicidae (Van Driesche & Bellows 1996).
Le nombre d'espèces de virus pathogènes aux insectes varie selon les auteurs de 650 (Cloutier & Cloutier 1992) à 1200 (Martignoni & Iwai 1981), la plupart (70%) ayant été décrits chez les Lépidoptères. On divise les virus entomopathogènes en 16 familles de virus à ADN ou ARN, à brins doubles ou simples. Deux familles seulement, les Baculoviridae et les Tetraviridae, n'infectent que des arthropodes, les autres familles ayant des représentants associés aux mammifères ou à d'autres groupes non-arthropodes, ce qui limite évidemment leur intérêt en lutte biologique.
Les 700 espèces de champignons entomopathogènes se regroupent dans cinq sous-divisions: les Mastigomycotina, Zygomycotina, Ascomycotina, Basidiomycotina et Deuteromycotina. Seulement environ une vingtaine d'espèces ont cependant été étudiées dans un contexte de lutte biologique.
Finalement les protozoaires infectant les insectes se retrouvent dans six phyla: les Zoomastigina (Flagellés), Rhizopoda (Amibes), Apicomplexa (Grégarines, Eugrégarines, Néogrégarines, Coccidies), Microspora (Microsporidies), Haplosporidia et Ciliophora (Ciliés). Pratiquement tous les travaux en lutte biologique ont porté sur les microsporidies.
Les nématodes entomophages exploitent les insectes comme ressource pour se développer et se reproduire. On retrouve des nématodes entomophages dans 30 familles différentes ce qui représente environ 4,000 espèces (Van Drische & Bellows 1996). La relation avec leur hôte va du simple commensalisme, sans aucun effet néfaste, jusqu'au parasitisme obligatoire, causant la mort de l'insecte. Dans cette dernière éventualité, les nématodes peuvent avoir un impact sur les populations de l'insecte hôte et ont suscité un intérêt en lutte biologique.
Parmi les principales familles où se retrouvent ces espèces, les Steinernematidae, Heterorhabditidae et Mermithidae sont celles offrant le meilleur potentiel en lutte biologique. Les genres ayant fait l'objet de plus d'études, les Steinernema et Heterorhabtidis, ne contiennent d'ailleurs que peu d'espèces, 10 et 3 respectivement (Doucet & Doucet 1990; Poinar 1990). Les nématodes entomophages s'attaquent à pratiquement tous les ordres d'insectes mais ce sont chez les Coléoptères, Diptères, Hyménoptères et Lépidoptères que le plus de cas ont été rapportés. Cependant cette prédominance pourrait bien ne refléter que la concentration des études dans ces groupes. Les prédateurs, qui tuent et consomment leurs proies (Price 1984) aux stades adulte ou larvaire, constituent un groupe beaucoup plus diversifié que les parasitoïdes. On retrouve des espèces prédatrices de façon importante chez neuf ordres d'insectes (Van Driesche & Bellows 1996) bien que ce soit chez les Hémiptères, Coléoptères, Diptères et Hyménoptères que la majorité de ces espèces se retrouvent.
 |
Figure 2- Femelle de Megarhyssa atrata (Hymenoptera: Ichneumonidae). Ces femelles percent le bois des arbres pour aller parasiter des larves de mouche à scie jusqu'à 15 cm à l'intérieur de l'arbre.
Chez les Hémiptères, plusieurs familles aquatiques sont prédatrices (Notonectidae, Pleidae, Naucoridae, Belostomatidae etc..). En plus de ces familles, les Anthocoridae, Nabidae, Reduviidae, Pentatomidae, Miridae, Phymatidae et Lygaeidae sont des prédateurs terrestres. Certaines de ces familles, dont les Miridae, contiennent aussi plusieurs espèces phytophages. Les Coléoptères contiennent probablement la majorité des espèces d'insectes prédateurs (Clausen 1940). Les familles de Coléoptères prédateurs les plus connues sont les Coccinellidae, Carabidae, Staphylinidae, Cicindelidae, Dytiscidae et Gyrinidae. Chez les Diptères, on retrouve des familles où seules quelques espèces sont prédatrices, tels les Tipulidae, Culicidae, Chironomidae, Rhagionidae, Asilidae, Empidae, Dolichopodidae, Drosophilidae et d'autres où la majorité des espèces ont un mode de vie de prédateur: Cecidomyiidae, Syrphidae et Chamaemyiidae.
Les parasitoïdes sont des organismes qui se développent sur ou dans un autre organisme, leur hôte, en tirent leur subsistance et le tuent comme résultat direct ou indirect de leur développement (Eggleton & Gaston 1990). Ces organismes ont une importance considérable autant d'un point de vue écologique que de lutte biologique. L'origine, la diversité et l'utilisation en lutte biologique de ces parasitoïdes feront l'objet des sections suivantes.
Les parasitoïdes
La première image qui nous vient à l'esprit lorsque l'on mentionne le mot "parasitoïde" est celle d'une guêpe parasitoïde (probablement d'ailleurs un Ichneumonidae ou un Braconidae) (Figure 2). Le monde des parasitoïdes est cependant beaucoup plus vaste et diversifié que ceux des guêpes de ces groupes. Les parasitoïdes appartenant aux Coleoptères (Figure 3) et Diptères sont aussi relativement bien connus, surtout par les spécialistes travaillant en lutte biologique, mais bien peu connaissent les parasitoïdes appartenant à des groupes comme les Lépidoptères et les Trichoptères. Le mode de vie parasitoïde, tel que défini plus haut, représente entre 5 et 20% des espèces d'insectes (Askew 1971, Eggleton & Belshaw 1992, Godfray 1994). Bien qu'il n'y aît approximativement que 800,000 espèces d'insectes décrites (Gaston 1991), l'évaluation du nombre total d'espèces d'insectes varie entre 2,5 et 30 millions d'espèces (Erwin 1983, Wilson 1988). Il pourrait donc y avoir de 125,000 à 6 millions d'espèces d'insectes ayant un mode de vie parasitoïde. On compte actuellement environ 87,000 espèces de parasitoïdes décrites dans le monde (Eggleton & Belshaw 1992) soit un peu plus de 10 % des espèces décrites. L'imprécision des évaluations tient beaucoup au fait que l'on ne connaît pas de façon précise la richesse de la faune entomologique de la plupart des milieux tropicaux. Par exemple on considère qu'au moins 75% des espèces d'Hyménoptères sont encore non-décrites et qu'une proportion importante des espèces décrites ne peuvent être identifiées (LaSalle 1993).
On retrouve des espèces ayant un mode de vie parasitoïde dans 6 ordres: Hyménoptère, Coléoptère, Diptère, Neuroptère, Lépidoptère et Trichoptère. Evidemment l'importance relative du mode de vie parasitoïde varie beaucoup d'un ordre à l'autre. Ainsi, les 67,000 espèces d'Hyménoptères parasitoïdes représentent environ 67% des espèces de cet ordre. C'est dans cet ordre que se retrouvent la plus grande diversité de modes de vie et d'adaptations à différents hôtes et habitats. L'ordre des Diptères compte un pourcentage important d'espèces parasitoïdes avec 16,000 espèces représentant 18% des espèces. L'importance du mode de vie parasitoïde est minime chez les autres ordres avec environ 1% des espèces parasitoïdes. Ainsi les Coléoptères ont 4,000 espèces de parasitoïdes sur 300,000 espèces, les Neuroptères 50 espèces sur 4500, les Lépidoptères 11 sur 113,000 et enfin les Trichoptères une espèce sur 4,500.
 |
Figure 3- Adulte de Aleochara bilineata (Coleoptera: Staphylinidae). Ces insectes sont des prédateurs d'oeufs de Diptères au stade adulte mais leur premier stade larvaire est un parasitoïde des pupes de ces mêmes Diptères.
Le nombre d'espèces de parasitoïdes dépasse donc certainement les 100,000 et ces espèces représentent une grande diversité. D'un point de vue de lutte biologique ce sont bien sûr des organismes d'une grande importance, mais quelle est leur importance dans les systèmes naturels? L'importance relative de chacun des groupes d'entomophages est variable. Lorsque les facteurs de mortalité biotique sont compilés pour 78 espèces d'herbivores ayant fait l'objet de tables de survie, les parasitoïdes ressortent comme le groupe causant la plus forte mortalité (Hawkins et al. 1997) (Figure 4). Il est aussi intéressant de noter que, globalement, le taux de mortalité biotique croit avec le stade de développement des herbivores (Figure 5). Ceci pourrait être dû à la faible rentabilité des oeufs et des jeunes stades larvaires pour les prédateurs. Cependant, une partie de cet effet provient sans doute de la difficulté d'identifier correctement les mortalités causées par des pathogènes ou des parasitoïdes tôt dans le développement d'un herbivore (Hawkins et al. 1997).
Il semble donc que, d'un point de vue écologique, la mortalité causée par les parasitoïdes soit la plus importante. La plus grande spécificité des parasitoïdes et leur capacité à localiser leurs hôtes peuvent expliquer cette plus grande importance. Enfin, les pathogènes, microorganismes et nématodes confondus, sont probablement limités par les conditions du milieu telles l'humidité et la lumière. Les épizooties causées par les pathogènes sont souvent spectaculaires mais peu fréquentes comparées au parasitisme qui est relativement constant d'une génération à l'autre.
Les Hyménoptères parasitoïdes sont nettement le groupe d'organisme le plus important en lutte biologique et il est responsable de la majorité des succès tant du point de vue économique qu'environnemental (LaSalle 1993). Globalement, depuis les 100 dernières années, environ 1200 introductions d'ennemis naturels, tous groupes confondus, ont été faites dans le monde. De ces introductions, 43% ont résulté en l'établissement de l'ennemi naturel et en une réduction au moins partielle des populations du ravageur visé (Van Driesche & Bellows 1996) mais les parasitoïdes se sont établis dans une proportion double de celle des prédateurs et ont été deux fois plus efficaces (Greathead 1986). Cependant, seulement 17% de ces introductions ont permis de contrôler efficacement le ravageur visé. Bien que ce pourcentage de succès apparaisse bas, il reste cependant que l'introduction d'ennemis naturels, lorsqu'elle est bien planifiée, est une approche ayant un rapport coût-bénéfice intéressant.
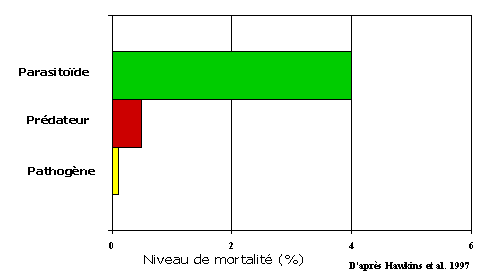 |
Figure 4- Mortalité naturelle d'insectes phytophages attribuée aux parasitoïdes, prédateurs et pathogènes. Données basées sur des tables de survie de 78 espèces d'herbivore
Les causes d'échecs rapportées par les auteurs varient selon que l'organisme introduit est un parasitoïde ou un prédateur. Alors que la race et l'absence d'hôtes alternatifs sont considérés comme des éléments importants dans le cas des parasitoïdes, ce sont plutôt la présence de refuges pour les proies et la compétition avec la faune indigène qui sont les plus souvent mentionnés pour les prédateurs (Figure 6) (Stiling 1993). La contrainte la plus importante demeure cependant le climat qui est considéré comme la cause d'échec dans le tiers des cas et ce autant pour les parasitoïdes que pour les prédateurs. Bien que possiblement biaisées parce que rapportées par les équipes ayant elles-même organisé les relâchers, ces explications permettent quand même d'espérer une amélioration dans le pourcentage de succès des introductions d'ennemis naturels exotiques. En effet, des programmes de sélection de parasitoïdes et de prédateurs permettraient de relâcher des souches mieux adaptées aux contraintes du climat et à la race du ravageur visé. Souvent, les raisons expliquant le choix d'une souche ont peu à voir avec la réalité du terrain. Ainsi, dans le cas du parasitoïde Trichogramma, une sélection rationnelle préalable n'est faite que dans 8.6% des cas où le choix de la souche est donné (Pak 1992). La souche utilisée a été choisie parce que facilement accessible ou abondante localement dans plus de la moitié des cas! La mise en place de protocoles standards permettant d'évaluer les caractéristiques biologiques des souches puis de choisir la souche la mieux adaptée au ravageur visé dans un environnement donné est certainement une des priorités en lutte biologique.
L'utilisation des parasitoïdes en lutte biologique offre plusieurs avantages. Ainsi on considère que les parasitoïdes ont une bonne capacité de dispersion et de découverte de l'hôte, ont une bonne capacité à s'établir dans un habitat donné, sont très sécuritaires pour la santé humaine et ont une grande spécificité d'hôte, donc posent peu de risques aux organismes non-visés (Cloutier & Cloutier 1992). Par contre, les parasitoïdes sont coûteux à élever en masse, sont fragiles, ce qui complique leur transport des sites de production vers les zones de relâcher, et les techniques de relâcher de parasitoïtes sont souvent compliquées et laborieuses. Il y a également un délai d'action entre le lâcher de parasitoïdes et l'effet recherché sur les hôtes. Leur grande spécificité, qui est un avantage lorsque ce sont les espèces non-visées qui sont considérées, devient un inconvénient économique puisque le développement d'une méthode de lutte biologique basée sur un parasitoïde doit se rentabiliser sur seulement un ou quelques hôtes (Cloutier & Cloutier 1992).
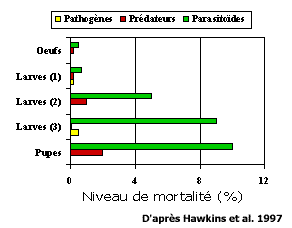 |
Figure 5- Mortalité naturelle d'insectes phytophages attribuée aux parasitoïdes, prédateurs et pathogènes selon le stade de développement de l'herbivore. Données basées sur des tables de survie de 78 espèces d'herbivore.
depuis quelques années, les introductions d'ennemis naturels sont réglementées et un permis d'introduction est exigé. Toute demande de permis d'introduction doit faire la preuve de la pertinence de cette introduction ainsi que des risques potentiels. Comme les prédateurs ont généralement un spectre d'hôte plus large que les parasitoïdes, les introductions de nouveaux prédateurs au pays sont particulièrement surveillées.
D'autres risques existent toutefois. La production de parasitoïdes tels les trichogrammes est basée sur une production de papillons hôtes en très grande quantité. Or les papillons ont les ailes couvertes de petites écailles et ces écailles se détachent tout au long de la vie d'un papillon. Les élevages de papillons entrainent donc de grandes concentrations d'écailles qui flottent dans l'air. Ces écailles peuvent causer des troubles respiratoires allant de la simple allergie à l'asthme et nécessitent de la part du personnel travaillant dans ces élevages le port de matériel respiratoire filtrant l'air (Brenner et al. 1991). Il s'agit donc dans ce cas d'une pollution reliée à la production des parasitoïdes mais qui est limitée aux locaux où se fait l'élevage.
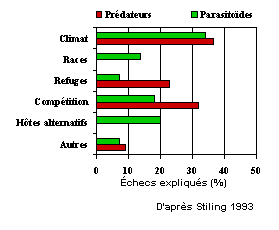 |
Figure 6- Causes d'échec dans des programmes de lutte biologique impliquant des parasitoïdes ou des prédateurs. Basé sur 126 cas (parasitoïdes) et 22 cas (prédateurs).
D'autres types de pollution sont plus insidieux. Ainsi la multiplication et le relâcher de grandes quantités d'un ennemi naturel indigène peuvent aussi causer une pollution dite génétique. Lorsqu'on débute un élevage de masse, on utilise un nombre réduit d'individus (de quelques dizaines à quelques milliers) mais cet échantillon ne représente nécessairement qu'une fraction de la diversité génétique d'une espèce. De plus, au cours d'un élevage de masse, la diversité génétique de l'élevage tendra à baisser, la sélection favorisant les individus pré-adaptés aux conditions de l'élevage. Il en résulte que, lors d'un relâcher, ce sont des individus génétiquement plus homogènes que les populations sauvages qui sont introduits en grand nombre dans l'environnement. Ces individus domineront l'habitat pendant une période de temps variable et durant cette période pourront s'accoupler avec les individus sauvages. Il y a alors un risque que des gènes maladaptés au milieu naturel soient introduits dans les populations sauvages. Par exemple, si les individus de l'élevage résistent mal aux basses températures, cette caractéristique ne sera jamais éliminée dans un élevage à température fixe. Cependant, si cette caractéristique est transmise à un grand nombre d'individus métissés (parents sauvage et d'élevage), ces individus et leurs descendants pourraient mal survivre à l'hiver. Les relâchers d'un programme de lutte biologique auraient alors comme résultat à moyen terme d'abaisser les populations sauvages du parasitoïde! L'importance de ce risque variera selon le niveau de variabilité génétique présent dans une espèce, caractéristique pour laquelle nous ne possédons à peu près pas d'information pour la grande majorité des ennemis naturels. Il serait important de mener des recherches dans ce domaine si on veut bien évaluer les bénéfices et les risques de la lutte biologique.
Il ressort donc que les parasitoïdes, et plus particulièrement les Hyménoptères, sont d'une grande importance autant dans les systèmes naturels qu'en lutte biologique. Malgré cette importance, les Hyménoptères parasitoïdes sont souvent présents en faible densité de population dans l'environnement. On pense que ces bas niveaux de population sont dus d'une part au fait que la plupart des Hyménoptères parasitoïdes sont relativement spécialisés et qu'ils agissent sur les populations de leurs hôtes d'une manière dépendante de la densité. En écologie, on qualifie de dépendant de la densité un facteur de mortalité qui augmente avec les populations de proies ou d'hôtes et qui inversement diminue lorsque ces populations diminuent. En conséquence, les niveaux d'équilibre des hôtes et des parasitoïdes sont relativement bas. Le fait que beaucoup d'espèces d'Hyménoptères parasitoïdes soient présentes en basse densité les rends susceptibles aux perturbations environnementales (LaSalle 1993). Or, parmi ces perturbations, plusieurs sont directement reliées aux activités humaines. L'utilisation des pesticides est évidememnt mise en cause mais la présence accrue de poussières dans les zones agricoles ou forestières peut diminuer le niveau de parasitisme (Rose & DeBach 1990). La fragmentation des milieux naturels, un problème important en milieu agricole, peut aussi contribuer à perturber l'équilibre fragile entre hôtes et parasitoïdes.
Malgré leur potentiel certain en lutte biologique, l'utilisation des parasitoïdes est freinée surtout par le manque d'information sur leur comportement et leur écologie. La production en masse de ces organismes et les relâchers efficaces dépendent largement de la qualité de notre compréhension du fonctionnement de ces organismes. Les recherches récentes dans ce domaine indiquent que dans un élevage de masse le nombre et la qualité des individus produits peut facilement varier du simple au double selon les conditions d'élevages. Or on ne connaît les conditions optimales d'élevage que pour quelques espèces de parasitoïdes. Des progrès considérables sont à prévoir dès que ces phénomènes seront mieux compris.
Remerciements
Je remercie Dominique Pelletier et Mathieu St-Louis pour leurs commentaires sur une version précédente de ce texte.
Bibliographie
Askew RR (1971) Parasitic insects. Heinemann Inc., London. Brenner RJ, Barnes KC, Helm RM, Williams LW (1991) Modernized society and allergies to arthropods. Am. Entomol. 37: 143-155.
Clausen CP (1940) Entomophagous insects. McGraw-Hill, New York.
Cloutier C, Cloutier C (1992) Les solutions biologiques de lutte pour la répression des insectes et acariens ravageurs des cultures. In: Vincent C & Coderre D (éd) La lutte biologique. p. 19-88. Gaëtan Morin Ed., Boucherville.
Doucet MMA, Doucet ME (1990) Steinernema ritteri (Nematoda: Steinernematidae) with a key to the species of the genus. Nematologia, 36: 257-265.
Eggleton P, Belshaw R, (1992) Insect parasitoids: an evolutionary overview. Phil. Trans. R. Soc. Lond. B 337: 1-20.
Eggleton P, Gaston KJ (1990) "Parasitoid" species and assemblages: convenient definitions or misleading compromises? Oikos 59: 417- 421.
Erwin TL (1983) Tropical forest canopies: the last biotic frontier. Bull. Entomol. Soc. Am. 29(1): 14-19.
Gaston KJ (1991) The magnitude of global insect species richness. Conserv. Biol. 5: 283-296.
Godfray HCJ (1994) Parasitoids, behavioral and evolutionary ecology. Princeton University Press, Princeton.
Greathead DJ (1986) Parasitoids in classical biological control In Waage J and Greathead DJ (éd) Insects parasitoids. p. 289-318. Academic Press, London.
Hawkins BA, Cornell HV, Hochberg ME (1997) Predators, parasitoids, and pathogens as mortality agents in phytophagous insect populations. Ecology 78: 2145-2152.
LaSalle J (1993) Hymenoptera and biodiversity In Lasalle J and Gauld ID (éd) Hymenoptera and biodiversity. p. 197-215, CAB International, Wallingford.
Martignoni ME, Iwai PJ (1981) A catalogue of viral diseases of insects, mites and ticks. In Burges HD (éd) Microbial control of pests and plant diseases, 1970-1980. p. 897-911. Academic Press, London.
Pak GA (1992) Inundative release of Trichogramma for the control of cruciferous Lepidoptera: preintroductory selection of an effective parasitoid. In Talekar NS (éd) Diamondback moth and other crucifer pests. pp. 297-308. Tainan, Taiwan : Asian Vegetable Research and Development Center, AVRDC Publication, No. 92-368.
Pedigo LP (1996) Entomology and pest management. 2nd ed. Prentice Hall, Upper Saddle River, New Jersey.
Poinar GO (1990) Taxonomy and biology of Steinernematidae and Heterorhabditidae. In Gaugler R & Kaya HK (éd) Entomopathogenic nematodes in biological control. p. 23-61. CRC Press, Boca Raton, Florida. Price PW (1984) Insect ecology. 2ième Edition, John Wiley & Sons, New York.
Rose M, DeBach P (1990) Foreign exploration and importation of natural enemies. In Rosen D (éd) Armored scale insects: their biology, natural enemies and control. p. 417-431. Elsevier, Amsterdam. Simberloff D, Stiling P (1996a) How risky is biological control? Ecology 77: 1965-1974.
Simberloff D, Stiling P (1996b) Risks of species introduced for biological control. Biol. Conserv. 78: 185-192.
Stiling P (1993) Why do natural enemies fail in classical biological control programs? Am. Entomol. 39: 31-37.
Van Driesche RG, Bellows TS (1996) Biological control. Chapman & Hall, New York
Wilson EO (1988) The current state of biological diversity, pp 3-18 in Wilson EO (Ed), Biodiversity, National Academy Press, Washington, D. C.
VertigO no 2, vol 2