1En Afrique équatoriale et tropicale, les formations ligneuses sont fortement sollicitées pour leur valeur commerciale ou médicinale et par la recherche de nouveaux espaces pour l’agriculture et l’élevage. Les écosystèmes forestiers denses ou clairs, de même que les arbres hors forêt dans les régions tropicales plus sèches, sont en voie de dégradation prononcée et très rapide depuis plus d’une cinquantaine d’années. Les liens entre la dégradation de l’environnement et les effets sur le développement durable de ces contrées sont maintenant bien connus (FAO 2005).
2En Ouganda, et particulièrement dans les zones habitées du Sud-Ouest du pays, les espèces en déclin comme Beilschmiedia ugandensis Rendle, Warburgia ugandensis Sprague (toutes deux figurant sur la liste rouge de l’IUCN), ou encore Zanthoxylum gilletii Waterman, Hallea rubrostipulata K Schum. et Spathodea campanulata Beauv., sont de plus en plus recherchées par les populations locales. Il est alors indispensable d’étudier leurs modes de régénération afin de proposer des solutions adéquates à leur conservation et de permettre leur réintroduction dans les milieux agro-pastoraux.
3La reproduction sexuée des ligneux est assez bien connue, mais ce mode de régénération nécessite un investissement matériel assez important (pépinière), ainsi qu’une bonne connaissance de la physiologie des semences (récolte, conditionnement, germination). Par contre, les études concernant la multiplication végétative à faible coût sont peu fréquentes pour les espèces tropicales (Bellefontaine 2005).
- 1 Le drageonnage est un procédé de propagation végétative permettant à certaines espèces, arborescen (...)
4Afin de proposer une méthode adéquate de multiplication peu coûteuse par induction du drageonnage1, il était indispensable de mieux comprendre le comportement des plantes qui montrent cette aptitude. C’est en général une réponse physiologique d’origine complexe, observée dans différentes situations (racines intactes, blessées ou sectionnées), à différentes périodes de l’année (saison sèche ou humide), propre à chaque espèce et parfois limitée à certains clones au sein d’une espèce donnée (CSFD, 2004 ; Radicati et al., 1994 ; Schier, 1978 ; Ryan et Bell, 1989). La compréhension et la maîtrise des réponses physiologiques après un stress pourraient s’avérer utiles pour ultérieurement développer la régénération à faible coût de ligneux, notamment pour des espèces dont la reproduction sexuée reste difficile ou coûteuse.
5Les observations décrites par la suite ont été réalisées dans la Province de Bushenyi en 2005. Elles ont été menées lors d’une étude en partenariat avec les tradipraticiens (ou guérisseurs) du centre de soin traditionnel de Rukararwe. L’objectif était de développer des techniques de multiplication efficaces et peu onéreuses sur des espèces médicinales recherchées, afin de fournir à la fois la pharmacopée et des revenus complémentaires pour les femmes. La station d’étude est située à proximité de l’équateur. Le climat est caractérisé par deux saisons sèches : l’une en février et l’autre en juin et juillet. La moyenne des précipitations annuelles est de 1 200 mm.
- 2 Distal : se trouvant vers l’extrémité en voie de croissance, loin de son point de fixation (Maro (...)
6A la suite d’un stress important subi par l’arbre-mère (coupe massive de racines), un pied de S. campanulata a réagi vigoureusement (Fig. 1). L’observation s’effectue à la lisière d’un espace pâturé. Cet arbre (environ 10 m de haut pour 40 cm de diamètre à 1,30 m) a été planté en bordure d’un champ. Une tranchée de 40 cm de profondeur a été creusée, vraisemblablement en 2004, à 2 m de l’arbre-mère. Elle a engendré le sectionnement complet de sept racines de diamètre variable. L’une d’entre-elles a un diamètre de 10 cm et les autres ont un diamètre compris entre 0,5 et 2 cm. Le couvert arboré est peu important ; les racines sectionnées et mises à nu par la tranchée sont exposées au soleil une grande partie de la journée.
- 3 Proximal : situé près du point de fixation à la plante (soit dans notre cas, près de la souche). (...)
7Les tiges adventives (c’est-à-dire les drageons dans un premier temps démunis de racine) se forment exclusivement sur les extrémités des racines déconnectées de l’arbre-mère (drageonnage distal). Dans ce cas, toutes les racines déconnectées, et quelque soit leur taille, ont produit au minimum un axe feuillé. La racine la plus grosse porte trois drageons (D1 à D3) situés juste en amont d’une ramification de la racine primaire. Les racines encore connectées à l’arbre-mère n’ont pas engendré de drageons proximaux3.
8Sur les neuf drageons présents, seul l’axe feuillé D7 (racine-mère de diamètre 0,8 cm) présente une néoformation de radicelles. Il est également le seul à être apparu juste à l’aplomb de la section de la racine. Dans les autres cas, l’apparition du drageon a lieu à une dizaine de centimètres de la section, et nettement plus loin sur la plus grosse racine.
Figure 1 : drageonnage d’un S. campanulata consécutif à la coupe de sept racines. Ouganda, juin 2005.

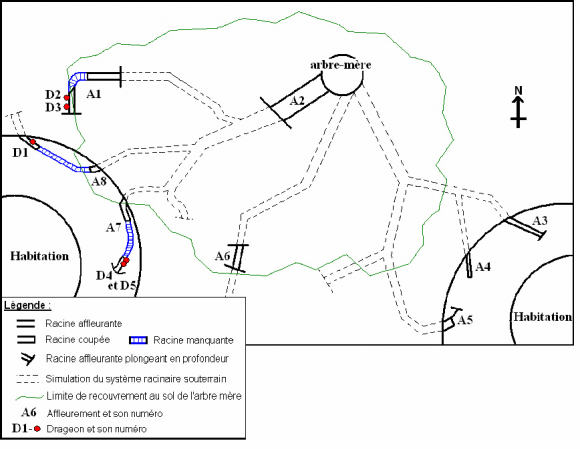
9Un cas similaire a été observé chez un autre S. campanulata dans un jardin. L’arbre-mère (environ 15 m de haut pour un diamètre de 38 cm à 1,3 m) est situé à égale distance de deux habitations rondes (Fig. 2). L’entretien régulier de la pelouse a engendré la mise à nu (affleurements) ou le sectionnement de plusieurs racines (A1 à A8). Trois de ces affleurements (A1, A7 et A8) ont donné naissance à cinq drageons (D1 à D5).
10Au total, huit affleurements ont été observés. Trois d’entre eux sont des racines coupées, d’un diamètre d’environ 3 cm, dont nous pouvons observer le prolongement quelques centimètres ou dizaines de centimètres plus loin (un tronçon de la racine est manquant). Dans les trois cas, la racine coupée (côté distal) et déconnectée de l’arbre-mère a produit un ou deux drageons (Fig. 2 et 3) et l’extrémité reliée à l’arbre-mère a produit des racines (radicelles dans le cas d’A8 et racine pivotante dans le cas d’A1) ou n’a manifesté aucune réaction apparente (A7).
11En A4 et A5, la racine a également été coupée (la partie déconnectée de l’arbre-mère n’a pas été retrouvée en 2005) et les extrémités connectées (proximales) ont produit de nouvelles racines pivotantes, mais pas de drageon.
12Enfin, pour les affleurements A2, A3 et A6, la racine est à l’air libre sur plusieurs dizaines de centimètres, blessée, mais non sectionnée. Dans ces trois cas, la racine n’a pas produit de drageon, ni de nouvelle racine.
13L’ombre produite par la cime de l’arbre-mère pourrait conditionner l’émergence des drageons, l’absence de lumière apparaissant comme un facteur limitant leur développement (Del Tredici, 1995). La position des cinq drageons, hors du recouvrement au sol de l'arbre –mère, confirme ce propos. Cependant, les affleurements A3, A4 et A5 (hors ombre) n’ont pas émis de drageon. La mise en lumière serait donc un facteur nécessaire mais non suffisant.
Figure 2 : schématisation des drageons et des affleurements racinaires chez S.campanulata. Ouganda, juin 2005.

14Ainsi, le sectionnement complet de la racine favorise la formation de drageons sur la partie déconnectée de l’arbre-mère (chez S. campanulata). Avant que l’axe feuillé du drageon n’apparaisse, ces racines déconnectées n’ont plus d’organes photosynthétiques et sont donc privées de toute alimentation en sève élaborée, sève permettant le maintien et la croissance des organes. La pérennité de ces segments de racine et de leurs tissus en milieu tropical chaud et humide est en principe très faible, mais l’émergence de ces axes feuillés permet la « survie des segments de racines isolées » et la réitération de nouvelles structures aériennes.
Figure 3 : apparition de deux drageons distaux (D4 et D5) suite au sectionnement d’une racine de S. campanulata, Ouganda, 2005.

15Une série d’expériences est menée en parallèle, de juin à août 2005. L’apparition de drageons succède en général à un stress subi par l’arbre-mère. Il s’agissait de tester les aptitudes au drageonnage de S. campanulata à la suite de blessures partielles du système racinaire. En blessant les racines d’un diamètre moyen de 1 à 3 cm, on suppose l’émergence d’un ou de plusieurs drageons au niveau de la blessure, voire en un autre point du système racinaire.
16Les expériences ont été effectuées sur quatre jeunes arbres. Au total onze racines superficielles ont été excavées, sur lesquelles ont été pratiquées:
17soit une annélation totale de la racine (sur 7 racines) en enlevant l’écorce tout autour de la racine sur 3 à 4 cm de long. (Fig. 4) ;
18soit une demi-annélation de la racine (sur 4 racines) en retirant l’écorce sur la moitié supérieure ou inférieure de la racine sur 3 à 4 cm de long.
19Pour ces onze expériences, la racine blessée a été immédiatement recouverte de la terre d’origine sur une épaisseur d’environ 5 cm.
Figure 4 : annélation complète en cours de réalisation sur une racine de S. campanulata

- 4 Le cal est une prolifération anarchique de cellules inorganisées, sans programme organogène, suiva (...)
20Après 90 jours, ces onze racines sont déterrées. Dans 91 % des cas (10 cas sur 11), on constate la formation d’un cal4 cicatriciel. Le cal prend naissance sur la partie proximale de la blessure et s’étend ensuite pour atteindre la partie distale. De ce fait, il recouvre la totalité de la blessure. La néoformation de racine est observée au niveau de la blessure lorsqu’il s’agit d’une annélation complète dans 3 cas sur 7 (Fig. 5).
Figure 5 : formation, après 90 jours, d’un cal cicatriciel et de racines au niveau d’une blessure infligée à une racine de S. campanulata.

21On constate sans surprise qu’une racine blessée sous terre produit un cal cicatriciel. Une fois la blessure recouverte entièrement par le cal, ce dernier tend à se durcir. A ce stade après 3 mois d’observation, la simple blessure de la racine, souterraine ou affleurante, n’engendre pas la formation d’un drageon.
22Les travaux réalisées sur S . campanulata en Ouganda ont été effectuées sur un nombre restreint d’individus du fait de leur faible densité dans la zone de l’étude. Les résultats n’ont pu être validés statistiquement. Cependant, ces premières observations sur six arbres mères et l’analyse du comportement des vingt-six racines révèlent une certaine uniformité des conditions d’apparition du drageon. Aussi, après trois mois d’observation, on peut mettre en avant les points suivants sur le drageonnage de cette espèce :
-
le sectionnement complet de la racine, laissée à l’air libre, engendre la formation d’axes feuillés distaux (drageons) souvent à quelques centimètres de la section ;
-
l’émergence du drageon distal est généralement accompagnée d’un renflement de l’axe primitif de la racine-mère, uniquement sur la partie déconnectée de l’arbre-mère ;
-
la partie racinaire reliée à l’arbre-mère forme de nouvelles radicelles sans émettre d’axe aérien (il n’y a pas formation d’un drageon) ;
-
l’affleurement de racines, suivi ou non de blessures (sévères ou légères) n’engendre pas de drageon ni de racines sur les racines-mères exposées ;
-
les racines souterraines blessées artificiellement « cicatrisent » par la formation d’un cal et peuvent induire la néoformation de racines, mais ne produisent pas de drageons en trois mois.
23L’émergence du drageon est post-traumatique. Il apparaît comme une réponse de survie des racines déconnectées de l’arbre-mère, qui, non alimentées, seraient vouées à disparaître. Les tiges adventives (c’est-à-dire les drageons sans racine dans un premier stade) ne sont pas issues d’une réponse directe de l’arbre-mère, mais bien de racines coupées et isolées. Apparemment, l’arbre-mère est capable d’engendrer deux types de structure au niveau de son système racinaire blessé :
24En revanche, l’arbre-mère semble incapable d’induire (toujours au niveau des racines blessées) une néoformation de tiges feuillées.
25Pourtant cette morphogenèse est systématique sur des racines déconnectées. Quel est le facteur déclenchant cette néoformation qui permet la survie d’un fragment de racine voué à disparaître ? Est-ce par le biais d’hormones, de flux nutritifs, ou encore de la redistribution des réserves glucidiques ? C’est autant de questions qui persistent au sujet des facteurs déclenchant ou inhibant cette organogenèse, et qui nécessitent de nouvelles observations et expérimentations, pour améliorer la connaissance de ces phénomènes.
26En Ouganda, des analyses similaires ont été menées parallèlement sur d’autres espèces. Dans des circonstances analogues (racines blessées, arbre-mère stressé), on observe des réponses physiologiques différentes selon l’espèce. On note par exemple un drageonnage exclusivement proximal (et non plus distal) chez Melia azedarach L., ou encore un drageonnage de type coloniaire (Hallé, 1999) chez Harungana madagascariensis Poir.. Le lieu de formation de drageons semble tout à fait propre à l’espèce.
27Si l’on veut utiliser le drageonnage comme outil de multiplication à faible coût, il est nécessaire de suivre la croissance et l’enracinement des drageons produits par induction. De nouvelles recherches consacrées aux conditions d’autonomie des drageons doivent compléter ces premières observations pour les espèces à valeur économique notable.
28Dans le contexte plus large du développement durable en Afrique et pour améliorer la santé des populations africaines rurales, il est indispensable de maintenir en vie la pharmacopée traditionnelle des villageois. Il est donc nécessaire de densifier le nombre d’arbres médicinaux à proximité de leurs maisons et des structures de soins de santé. Tous les moyens économiques de régénération (semis, boutures de tiges ou de racines, rejets de souche, drageons, marcottes) doivent être mobilisés en Afrique Tropicale et Equatoriale afin d’améliorer les conditions de vie locale et réduire la pauvreté, conditions sine qua non pour un développement durable. Les arbres ont une incidence directe sur les revenus complémentaires des femmes africaines, chargées notamment de la santé et de l’alimentation des familles. A la suite d’une enquête réalisée en 2005 avec 110 agropasteurs (Meunier, 2005), S. campanulata a été classée 5ème priorité parmi les 37 espèces mentionnées. Elle est vivement recherchée par la population locale, pour ses vertus médicinales notamment.
29Les descriptions chez S. campanulata pourront permettre d’anticiper et de contrôler l’apparition de drageons (dans l’espace et dans le temps) lors d’inductions artificielles.Cette étude de cas concrets nous permet d’envisager des solutions peu coûteuses pour multiplier des arbres à usages multiples, dont ceux qui sont nécessaires aux tradipraticiens. Dans cette perspective, H. rubrostipulata, W. ugandensis, B. ugandensis et Z. gilletiinécessitent une attention particulière. Recherchées par les populations rurales pour leur haute valeur médicinale, elles présentent des taux de germination très faibles, quasi inexistants en milieu naturel. La régénération asexuée doit être testée pour palier leur disparition prévisible.
30En associant et comparant les différentes techniques de régénération de chaque espèce, on pourra privilégier dans certains cas des méthodes de multiplication à faible coût pour une espèce donnée, et ainsi encourager les initiatives de maintien voire d’extension du patrimoine floristique. Il est nécessaire d’analyser méticuleusement les cas de drageonnage naturel avant d’envisager la propagation de ligneux par induction. Il faut étudier ces phénomènes à l’échelle de l’espèce, car il ne semble pas exister de méthode d’induction unique du drageonnage, chaque espèce présentant des réponses physiologiques propres. En Afrique, le développement durable passe nécessairement par la régénération à faible coût des ressources ligneuses, dont les populations souvent démunies, ont un cruel besoin.






{kind=link}