|
||||
MATERIALS AND METHODS
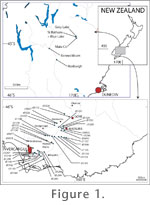
The 58 samples studied here come from outcrops of the St Bathans Member, in the old gold-sluicing areas of Blue Lake (sample numbers prefixed with "BL"), Grey Lake, about 2 km to the northeast (sample numbers prefixed with "GL"), and the bed of Mata Creek, about 3.5 km to the southeast of St Bathans (sample numbers prefixed with "Mata"). The fossils come from lenses of carbonaceous mud separated by quartzose gravels and probably all lay within a "St Bathans Paleovalley," which had been incised into the greywacke basement rock (Douglas 1986). The palynology of the St Bathans Member indicates an Early Miocene age (Mildenhall 1989; Mildenhall and Pocknall 1989), although perhaps an extension into the Oligocene cannot be ruled out. Grid references to all samples are given in Appendix 1. Eight-digit grid references preceded by H41 (the map number) were estimated from field observations and are based on the New Zealand 1:50 000 Topographic Map 260 series. Fourteen-digit references were made with a Garmin Geko 301 GPS unit and are full coordinates in terms of the New Zealand metric map grid (UTM zone 59). A realistic accuracy for these figures would be around 10-15 horizontal metres. The stratigraphy within the St Bathans Member is complicated by the usual channeling in a fluvial environment and is the subject of ongoing research. At this stage brief comments are given which show the relative stratigraphic difference between nearby samples, or their estimated height above basement rock. Where available, New Zealand Geological Society Fossil Record numbers are given in brackets in the Stratigraphy column of the Appendix. Further to the south, the lithostratigraphy of the Gore Lignite Measures has been documented by Isaac and Lindqvist (1990), who interpreted the unit as a large coastal delta and possibly represented the drainage point for the Manuherikia Group. The fossils have been obtained from a range of drill cores drilled as part of lignite exploration. The coal measures overlie and are intercalated with marine sediments. The associated marine invertebrates have facilitated their dating, which includes latest Oligocene and Early Miocene strata (Pocknall and Mildenhall 1984). The 62 samples studied here (prefixed by "Sthd-") all come from levels dated as Early Miocene. The location of all samples is given in Appendix 2. The drillcore number follows Isaac and Lindqvist (1990), and the location coordinates are taken directly from that publication (which should be consulted for the individual accuracy of these figures which varies from the nearest 0.1 of a metre to the nearest hundred metres). Sample depth is measured down from the top of the core. Most St Bathans Member samples consisted of about 300 g with about half of that amount being prepared. However, seven samples were more intensively sampled by repeat visits as they have more abundant and intact angiosperm fossils. These samples are BL-30, GL-01, GL-02, GL-32, H41/f038, BL-32, Mata-01 and up to one or two kilos from them will have been processed. The size of Gore Lignite Measure samples was significantly smaller than those from the St Bathans Member as they were limited by the diameter of the drill core (c. 50 mm) and the need to preserve the integrity of the core for future research. Sample size was about 50 g, and the amount of sediment prepared was about half of that. Sediment samples were broken down using boiling water and hydrogen peroxide, sieving, and removal of silicates with hydrofluoric acid. Intact specimens of mummified leaves are removed at this point (if more clearing is needed, they may be again placed in hydrogen peroxide) and mounted within sheets of plastic using thymol glycerine jelly. Clean cuticle is prepared by warming for several hours in aqueous chromium trioxide, washing, then staining with Crystal violet, and finally mounting on glass slides with thymol glycerine jelly for transmitted light microscopy (TLM). Where sufficient additional material existed, it was mounted on SEM (scanning electron microscope) stubs with double-sided cellotape and coating with platinum. For the sake of brevity, with one exception, only the stomatal (abaxial) surfaces are illustrated. In addition, at a few localities in the St Bathans Paleovalley intact leaves could be collected adhering to slabs of mudstone, or even gathered as they fell out when mudstone blocks were split. These specimens were also cleaned in hydrofluoric acid and gently cleared in weak hydrogen peroxide solution until the venation was visible. They were then mounted between sheets of clear plastic using glycerine jelly. As discussed above, extant Australasian Lauraceae cuticle has a distinctive structure whereby the guard cells are typically overarched by a pair of subsidiary cells (a paracytic structure), and a cuticularised flange develops between the two cell types (Hill 1986). Trichome bases, when present, are simple and poral. If fossil cuticle has this structure, it is assumed here to be Lauraceae. However, in some taxa the guard cells are not below the subsidiary cells, but adjacent, for example Endiandra (see Christophel and Rowett 1996, Plate 2C). There are extant taxa that have a somewhat similar structure and might be confused with Lauraceae in the fossil record, for example, Hedycarya (Monimiaceae). Four of the fossil taxa described below are glabrous, so their trichome bases structure is unknown. This is not unusual amongst extant Lauraceae (at least on small fragments), and the small possibility that a few of the taxa described here do not belong in the Lauraceae would not seriously effect the conclusions of this paper. Arriving at a practical conclusion to distinguish fragments of Lauraceae cuticle and consistently grouping them in over 120 samples has been the single most difficult job in this overall project (mostly due to subtle differences in morphology, such as the density of trichome bases preservation, and degree of staining). It is likely that some taxa distinguished here may include several closely related forms. As a result, overall diversity is likely to be underestimated. A key to the fossil cuticle is given in Appendix 3, and all descriptive taxonomy is given as Appendix 4. To keep morphological similar taxa together, the taxa are presented in the same order as found in the key. Leaf architecture and epidermal terminology follows Dilcher (1974); Hickey (1973); Hill (1986); and Pole (1991). The taxonomic method I have adopted is to give each recognisable cuticle morphology and leaf morphology a parataxon code. As these parataxa are not Latin binomials they are entirely outside of the ICBN. The ICBN recognizes 'morphotaxa', but as these are Latin binomials, the term is not used here. My concept of parataxa is that they are essentially species for purposes of biodiversity, but nomenclatural they are like Linnaean species without the genus. Dispersed Lauraceae cuticle taxa are named with the prefix "CUT-L-" followed by a unique string of three letters (see Carpenter and Pole 1995; Pole 1996, 1998, for examples of cuticle parataxon use). This gives some flexibility to a situation where there is little obvious hierarchy. Following Pole (1993b, and references therein) Manuherikia Group leaf parataxa are given the prefix "MANU-". For parataxa a reference specimen and reference locality is nominated, equivalent to holotype and type locality. A single "referred specimen" is also nominated from each of the other samples in which the taxon occurs. Having separate parataxa for leaves and cuticle recognises that leaves (as impressions with only gross morphology and vein architecture) and cuticle (as dispersed fragments) are commonly encountered separately in the geological record. I reject the common paleobotanical practice of developing a system of Linnaean binomials for these fragments which overtly look comparable to 'true' Linnaean taxa and yet in many cases seem to be little more than a restatement of the family to which the fragment belongs (e.g., Laurophyllum, which has been used in the past). In the few cases where identification with a Linnaean genus is possible, the Linnaean name is applied in addition to the parataxon code. The prefixes "OU," "SB," and "SL" refer to specimens mounted on glass slides or between sheets of plastic. Scanning Electron Microscope stubs are prefixed with "S-" . All material is housed in the Centre for Marine Studies, University of Queensland, except those prefixed by "OU," which are housed in the Geology Museum of the Otago University. |
||||
|
|