MICROFACIES CHARACTERISTICS AND SINTER PROPERTIES
Microfacies 1 – Cup- to Ridge-Shaped Sinter
 Microfacies
1 (Figure 3) sinter forms in
slightly turbulent waters with high emissions of steam, and water
temperatures ranging from 91°C at some vents to 64°C in the associated
discharge channels (Figure 4.1). The
water turbulence is caused by the ebullient discharge from nearby vents that
produces pulses of thermal water (typically 85°C) washing onto the siliceous
sinters. Neither splash nor spray was observed around these deposits.
However, water level changes were recorded in this microfacies (Figure
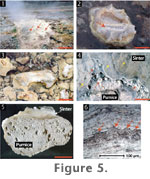
4.1). Sinter forms subaerially on pumice clasts and pine cones that act
as substrates above water level (Figure 5).
Sinter accretion rates were fastest at this microfacies. An average of 0.24
g silica accumulated on glass slides over 67 days in this environment (Figure
4). It is unknown if Microfacies 1 sinter accreted at a uniform
rate or not.
Microfacies
1 (Figure 3) sinter forms in
slightly turbulent waters with high emissions of steam, and water
temperatures ranging from 91°C at some vents to 64°C in the associated
discharge channels (Figure 4.1). The
water turbulence is caused by the ebullient discharge from nearby vents that
produces pulses of thermal water (typically 85°C) washing onto the siliceous
sinters. Neither splash nor spray was observed around these deposits.
However, water level changes were recorded in this microfacies (Figure
4.1). Sinter forms subaerially on pumice clasts and pine cones that act
as substrates above water level (Figure 5).
Sinter accretion rates were fastest at this microfacies. An average of 0.24
g silica accumulated on glass slides over 67 days in this environment (Figure
4). It is unknown if Microfacies 1 sinter accreted at a uniform
rate or not.
XRPD analyses of sinters from this and all
other microfacies of this study site showed that they are composed almost
entirely of opal-A, with minor traces of detrital quartz and/or feldspar
(for full width at half maximum intensity (FWHM) values and density/porosity
data, see Rodgers et al. 2004). DTA of sinters from this study also gave
identical traces characteristic of opal-A, with no indications of clay
content (cf. Herdianita et al. 2000a). TGA of these sinters indicated weight
loss of approximately 5 to 6% resulting from opal-A dehydration (cf.
Herdianita et al. 2000b; Handley et al. 2005).
Microfacies 1 sinter is vitreous, light
grey in colour, and usually cup- or ridge-shaped with minute microspicules
(0.5 cm high) visible on the uppermost sides of the rim or within cavities (Figure
5.2). The sinter surfaces have multiple, thin layers of silica with an
irregular texture, and are composed of gnarled, broken, and isolated surface
remnants (Figure 5.4). This irregular
texture is gradational, occurring predominantly in the lowermost portions of
the sinter, close to the air-water interface. Vertical sections through the
sinters exhibit alternating series of light brown and light grey laminae (Figure
5.5). The laminae range from <2 to 50 µm in thickness and alternate
between brown to dark brown and clear, translucent silica (Figure
5.6). No evidence for the presence of microorganisms or any
mineral-microbe association was observed in thin-section. Laminae in the
lower portions of the sinters are relatively flat, but become progressively
more convex upwards. On and within these layers, conical microspicules may
be present, but in a relatively lower frequency than in Microfacies 2. Under
SEM, vertical sections appeared vitreous and massive, without any
conspicuous laminae. Prokaryote-sized microorganisms were only rarely
observed in these sections.
At the Parariki site, sinter morphology is
affected by the subaerially exposed substrate dimensions. Substrates that
are relatively wider and higher above water exhibit limited siliceous
coating. This coating is confined to the outer fringes of the substrates,
resulting in the formation of cup-shaped deposits (Figure
5.2). On substrates that are low-lying and less aerially extensive,
silica deposited across the substrate and a ridge-shaped deposit arises (Figure
5.3).
 Large
areas of Microfacies 1 sinter surfaces are covered by irregularly
lobed, coccoidal microorganisms (1-1.5 µm in diameter) (Figure
6.1). These microbes are linked to one another through a meshwork of
mucosal to filamentous extracellular polymeric substances (EPS). Such
microbe-EPS assemblages are known as biofilms (cf. Cady and Farmer 1996; Handley et al. 2005). Biofilms in the upper- and inner-most portions of the
Parariki sinters typically assumed the shapes of isolated, conical cell
clusters (Figure 6.2). Some of these
clusters became progressively encrusted by nanospheres of silica (<250 nm in
diameter) and eventually recolonised by a new succession of microbes (Figure
6.3-6.4). Note that individual nanospheres attached directly to the
cells and the associated EPS, thereby starting the process of silicification.
Cells that appeared free from these silica spheres may be tentatively
regarded as unsilicified.
Large
areas of Microfacies 1 sinter surfaces are covered by irregularly
lobed, coccoidal microorganisms (1-1.5 µm in diameter) (Figure
6.1). These microbes are linked to one another through a meshwork of
mucosal to filamentous extracellular polymeric substances (EPS). Such
microbe-EPS assemblages are known as biofilms (cf. Cady and Farmer 1996; Handley et al. 2005). Biofilms in the upper- and inner-most portions of the
Parariki sinters typically assumed the shapes of isolated, conical cell
clusters (Figure 6.2). Some of these
clusters became progressively encrusted by nanospheres of silica (<250 nm in
diameter) and eventually recolonised by a new succession of microbes (Figure
6.3-6.4). Note that individual nanospheres attached directly to the
cells and the associated EPS, thereby starting the process of silicification.
Cells that appeared free from these silica spheres may be tentatively
regarded as unsilicified.
 The
continuous interplay between microcolony formation, silicification, and
recolonisation appears to have formed vertically upright, pillar-like
microspicules (Figure 7). The upper
surfaces of these spicules displayed a knobby morphology (typically 1-2 µm
in diameter) of similar dimensions to nearby coccoidal cells (Figure
7.3). Thus we infer that individual knobs formed by the silicification
of these microbial cells. The knobby texture was only observed on spicules
of Microfacies 1, where it coincided with the presence of coccoidal
microbial cells. We did not observe this texture on spicules from other
microfacies where bacilli (rod-shapes) predominate (see below).
The
continuous interplay between microcolony formation, silicification, and
recolonisation appears to have formed vertically upright, pillar-like
microspicules (Figure 7). The upper
surfaces of these spicules displayed a knobby morphology (typically 1-2 µm
in diameter) of similar dimensions to nearby coccoidal cells (Figure
7.3). Thus we infer that individual knobs formed by the silicification
of these microbial cells. The knobby texture was only observed on spicules
of Microfacies 1, where it coincided with the presence of coccoidal
microbial cells. We did not observe this texture on spicules from other
microfacies where bacilli (rod-shapes) predominate (see below).
The irregularly lobed cocci are regarded as
microbial cells and not as abiogenic crystals, since they are highly
irregular in shape, often associated with copious EPS (Figure
6) and are very similar in morphology under SEM to cultured archaeal
thermoacidophiles (e.g., Chen et al. 2005, figure 5A). Indeed, 16S rRNA
genes extracted from Microfacies 1 sinter is related to known
thermoacidophiles that also have irregularly lobed sphere shapes (e.g.,
Thermoplasmatales; Schinteie 2005).
 SEM
examination revealed that silica deposited predominantly as spheres of
opal-A that are primarily <10 nm to 50 nm in diameter. However, spheres can
grow up to 500 nm in diameter through the aggregation of smaller spheres.
Two types of sphere shapes were observed: (1) freshly deposited, round
equidimensional spheres; and (2) more poorly defined spheres with
interparticle "necks" (Figure 8.1-8.2)
(cf. Iler 1979, figure 3.24). The poorly defined sphere shape-texture was
pervasive on Microfacies 1 sinter. Associations between this texture
and microorganisms were not observed under SEM. In addition to silica, well
developed crystals of gypsum, barite, and sulphur were present. No clay
minerals were observed anywhere upon or within sinter from this or any other
microfacies at the Parariki site. The absence of clay minerals is in
contrast to acid-derived sinters and residues reported elsewhere in New
Zealand (Jones et al. 2000; Rodgers et al. 2002).
SEM
examination revealed that silica deposited predominantly as spheres of
opal-A that are primarily <10 nm to 50 nm in diameter. However, spheres can
grow up to 500 nm in diameter through the aggregation of smaller spheres.
Two types of sphere shapes were observed: (1) freshly deposited, round
equidimensional spheres; and (2) more poorly defined spheres with
interparticle "necks" (Figure 8.1-8.2)
(cf. Iler 1979, figure 3.24). The poorly defined sphere shape-texture was
pervasive on Microfacies 1 sinter. Associations between this texture
and microorganisms were not observed under SEM. In addition to silica, well
developed crystals of gypsum, barite, and sulphur were present. No clay
minerals were observed anywhere upon or within sinter from this or any other
microfacies at the Parariki site. The absence of clay minerals is in
contrast to acid-derived sinters and residues reported elsewhere in New
Zealand (Jones et al. 2000; Rodgers et al. 2002).
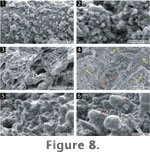
At the microscale, the surfaces of the lower
portions of Microfacies 1 sinter are irregular and uneven.
Anastomosing ridges, nodules, and associated cavities are common. Incipient
ridges appear as surface irregularities that are bounded by small cavities (Figure
8.3-8.4). Within these cavities, nodules formed that eventually become
mushroom-shaped, producing a neck-like structure (Figure
8.4-8.6). While incipient nodules are surrounded by small cavities,
taller nodules are associated with deeper cavities and larger ridges.
 The
lower portions of Microfacies 1 sinters exhibit isolated patches of
surficial silica sheets (Figure 9.1).
These sheets appeared to have become progressively contracted and
diminished, particularly around necks (Figure
9.2), so as to leave behind isolated remnants of a formerly continuous
surface. The patches are bound by anastomosing ridges (Figure
9.3) that are larger towards the outer fringes of these isolated patches
(Figure 9.4).
Figure 9.1-9.4 show the surface of
silica grown onto a glass slide for one month in a Microfacies 1
setting. Surfaces of natural Microfacies 1 sinter show even more
extreme forms of isolated remnants (Figure
9.5-9.6), presumably because they have been in this environment for much
longer than the slides. These remnants contain concentric laminae, mirroring
the topography of the underlying layers, as well as ridges, which are larger
on the fringes.
The
lower portions of Microfacies 1 sinters exhibit isolated patches of
surficial silica sheets (Figure 9.1).
These sheets appeared to have become progressively contracted and
diminished, particularly around necks (Figure
9.2), so as to leave behind isolated remnants of a formerly continuous
surface. The patches are bound by anastomosing ridges (Figure
9.3) that are larger towards the outer fringes of these isolated patches
(Figure 9.4).
Figure 9.1-9.4 show the surface of
silica grown onto a glass slide for one month in a Microfacies 1
setting. Surfaces of natural Microfacies 1 sinter show even more
extreme forms of isolated remnants (Figure
9.5-9.6), presumably because they have been in this environment for much
longer than the slides. These remnants contain concentric laminae, mirroring
the topography of the underlying layers, as well as ridges, which are larger
on the fringes.
 Microfacies
2 (Figure 3) is characterised by
quiescent thermal water discharge. Steam emission is also minor, while water
temperatures range from ~85°C to ~30°C (Figure
4.1). As in Microfacies 1, neither splash nor spray was observed
around these deposits. Microfacies 2 sinter forms subaerially on
pumice clasts, wood, pine cones, and dead insects (Figure
10). Water level changes have also been observed in this microfacies (Figure
4.1). Steam condensate keeps the sinters moist. Silica accretion rates
are slower than those in Microfacies 1; an average of 0.12 g silica
accreted on the slides over 67 days (Figure
4).
Microfacies
2 (Figure 3) is characterised by
quiescent thermal water discharge. Steam emission is also minor, while water
temperatures range from ~85°C to ~30°C (Figure
4.1). As in Microfacies 1, neither splash nor spray was observed
around these deposits. Microfacies 2 sinter forms subaerially on
pumice clasts, wood, pine cones, and dead insects (Figure
10). Water level changes have also been observed in this microfacies (Figure
4.1). Steam condensate keeps the sinters moist. Silica accretion rates
are slower than those in Microfacies 1; an average of 0.12 g silica
accreted on the slides over 67 days (Figure
4).
The outer and lower portions of
Microfacies 2 sinters are usually covered by a ubiquitous green
microbial mat (Figure 10).
Transmission electron microscopy (TEM) and rRNA gene analysis indicates that
these mats are primarily composed of coccoidal algal cells related to the
rhodophyte taxon Cyanidiophyceae (Cyanidium and Galdieria;
Schinteie 2005). The mats occur where temperatures are between 52.5 and
30°C. On sinter, the mats are at their thickest (1 mm) at ~ 45°C. On the
uppermost portions of the sinters, where temperatures are often slightly
cooler and less exposed to thermal water, the mats become faint and
disappear.
The sinters are white to vitreous in colour,
and typified by significant spicular growth (Figure
10). The spicules are needle-like to conical in shape, 1 to 3 mm wide,
and from <1 mm to ~1 cm long. Spicules are densely packed, accrete
perpendicular to the substratum, and become progressively smaller outward
towards the water. A rim forms where spicules progressively link laterally
with each other through a continuous deposition of silica (Figure
10.2, 10.5; cf. Handley 2004). Vertical sections through these sinters
revealed alternating light and dark laminae (Figure
10.6).
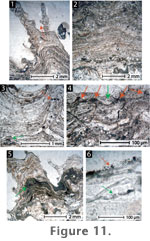 The
laminae alternate among laterally continuous green, brown, and translucent
silica (Figure 11.1). As in
Microfacies 1, laminae in the lower sinter portions are relatively flat
but become progressively convex upwards (Figure
11.2), culminating in spicules composed of parabolic laminae. The fate
of incipient spicules varied over time; some became reinforced by the
deposition of successive laminae, whereas others were smothered or dampened
by the lamination process (Figure
11.3-11.4). Overall, the relief of the sinters increased over time, with
surfaces characterised by abundant, erect, and free-standing spicular
structures with parabolic laminae. Numerous spicules also exhibit branching
and small (~0.5 mm) projections with internal convex laminae.
The
laminae alternate among laterally continuous green, brown, and translucent
silica (Figure 11.1). As in
Microfacies 1, laminae in the lower sinter portions are relatively flat
but become progressively convex upwards (Figure
11.2), culminating in spicules composed of parabolic laminae. The fate
of incipient spicules varied over time; some became reinforced by the
deposition of successive laminae, whereas others were smothered or dampened
by the lamination process (Figure
11.3-11.4). Overall, the relief of the sinters increased over time, with
surfaces characterised by abundant, erect, and free-standing spicular
structures with parabolic laminae. Numerous spicules also exhibit branching
and small (~0.5 mm) projections with internal convex laminae.
The green laminae observed in thin section (Figure
11.5) are composed of spheres that are similar in size (2-10 µm) and
appearance to the coccoidal cells present in the green biomats (Figure
11.6). These cells are restricted to the lower and middle portions of
Microfacies 2 sinter, corresponding to the distribution of the living
green mats. Diatom tests occur throughout the sinter.
As in Microfacies 1, sinter
morphology also is controlled by substrate shape and dimensions. Subaerial
substrate portions that are relatively wider and higher above water exhibit
less siliceous encrustation relative to substrate size than deposits that
are low-lying and have smaller widths. While wider substrates display sinter
with cup-shaped morphologies (Figure
10.2), smaller substrates are covered in spicules that coat much of the
subaerial portions (Figure 10.3).
The extent of siliceous covering in Microfacies 2 is less than that
of comparable substrate sizes and shapes in Microfacies 1, where
relatively higher energy conditions occur.
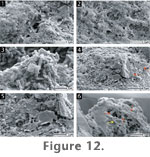 Bacilli
(i.e., rod-shaped microorganisms) (1-2.3 µm long) dominate the sinter biota
of this microfacies (Figure 12.1).
The microbes are usually associated with a meshwork of EPS that includes
both fibrous and mucosal textures. These biofilms became progressively
silicified by coalesced to partially coalesced opal-A nanospheres (100 nm)
and microspheres (250 nm) (Figure 12.2).
Eventually, the films were completely obliterated by the deposition of
overlying spheres.
Bacilli
(i.e., rod-shaped microorganisms) (1-2.3 µm long) dominate the sinter biota
of this microfacies (Figure 12.1).
The microbes are usually associated with a meshwork of EPS that includes
both fibrous and mucosal textures. These biofilms became progressively
silicified by coalesced to partially coalesced opal-A nanospheres (100 nm)
and microspheres (250 nm) (Figure 12.2).
Eventually, the films were completely obliterated by the deposition of
overlying spheres.
The bacilli also tended to form clumps of
microcolonies which grew perpendicular to the silica surface and were
associated with a network of EPS (Figure
12.3). In places, diatom tests provided areas of positive relief onto
which new clusters of bacilli aggregated. As in Microfacies 1, such
microcolonies became silicified, forming areas of positive relief (Figure
12.4). This consistent interplay between bacterial colonisation and
silicification also resulted in the formation of microspicules. The possible
outlines of bacilli inside spicules are preserved (Figure
12.6). The occurrence of vertically upright microcolonies in
Microfacies 2 is more widespread than in Microfacies 1.
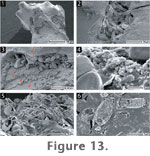 Pennate
diatoms, predominantly Pinnularia acoricola Hustedt and P.
champmaniana Foged, constitute a major component of the sinter biota (Schinteie
2005; cf. Foged 1979; Cassie 1989; Cassie and Cooper 1989). These diatoms
preferentially occupied areas of low microrelief, such as crevices and
cracks, as well as overlying areas of positive relief (Figure
13.1-13.3). The mode of attachment for these benthic diatoms is adnate,
or closely appressed to the substratum, with the entire valve attached to a
substrate by a coating of EPS (Figure
13.4). In open areas of the deposits that do not offer sheltering by
surrounding sinter, diatom tests were often found fractured and amassed into
clumps (Figure 13.5). Diatom
assemblages may eventually become part of the sinter deposit, starting with
the precipitation of silica spheres onto the tests, which eventually result
in their complete cementation (Figure
13.6) (cf. Jones et al. 2000; Campbell et al. 2004). In this and in
other microfacies of the Parariki site, early silicification occurs
preferentially on the edges of diatom tests, EPS sheets, and fibres.
Pennate
diatoms, predominantly Pinnularia acoricola Hustedt and P.
champmaniana Foged, constitute a major component of the sinter biota (Schinteie
2005; cf. Foged 1979; Cassie 1989; Cassie and Cooper 1989). These diatoms
preferentially occupied areas of low microrelief, such as crevices and
cracks, as well as overlying areas of positive relief (Figure
13.1-13.3). The mode of attachment for these benthic diatoms is adnate,
or closely appressed to the substratum, with the entire valve attached to a
substrate by a coating of EPS (Figure
13.4). In open areas of the deposits that do not offer sheltering by
surrounding sinter, diatom tests were often found fractured and amassed into
clumps (Figure 13.5). Diatom
assemblages may eventually become part of the sinter deposit, starting with
the precipitation of silica spheres onto the tests, which eventually result
in their complete cementation (Figure
13.6) (cf. Jones et al. 2000; Campbell et al. 2004). In this and in
other microfacies of the Parariki site, early silicification occurs
preferentially on the edges of diatom tests, EPS sheets, and fibres.
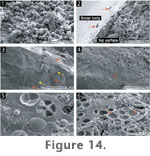 The
green algal-dominated mats present on the lower surfaces and in thin section
of Microfacies 2 sinters are largely composed of colonies of
spherical cells (2-10 µm in diameter) that are covered in membranous sheets
(Figure 14.1). The cell surfaces of
these mats can become encrusted and incorporated into lower portions of the
sinter (Figure 14.2-14.4; cf.
Figure 11.5-11.6). Vertical sections
through these portions exhibit numerous horizontal laminae (~100-180 µm
thick) of dense, vitreous sinter, alternating with layers (~80-120 µm thick)
dominated by silicified cells that are equal in size to cells of the living
green mats (Figure 14.5). Several
silicified cells also exhibited endospores (i.e., internal division of
parental algal cells). In some instances, cellular impressions were
preserved (Figure 14.6). The
boundary between the predominantly abiotic and biotic layers is sharp and
planar, showing that cells of the green mats initially colonised a flat
surface before becoming silicified (Figure
14.4).
The
green algal-dominated mats present on the lower surfaces and in thin section
of Microfacies 2 sinters are largely composed of colonies of
spherical cells (2-10 µm in diameter) that are covered in membranous sheets
(Figure 14.1). The cell surfaces of
these mats can become encrusted and incorporated into lower portions of the
sinter (Figure 14.2-14.4; cf.
Figure 11.5-11.6). Vertical sections
through these portions exhibit numerous horizontal laminae (~100-180 µm
thick) of dense, vitreous sinter, alternating with layers (~80-120 µm thick)
dominated by silicified cells that are equal in size to cells of the living
green mats (Figure 14.5). Several
silicified cells also exhibited endospores (i.e., internal division of
parental algal cells). In some instances, cellular impressions were
preserved (Figure 14.6). The
boundary between the predominantly abiotic and biotic layers is sharp and
planar, showing that cells of the green mats initially colonised a flat
surface before becoming silicified (Figure
14.4).
Unlike sinter surfaces of Microfacies 1,
those from Microfacies 2-4 are generally not irregular in appearance.
Ridges, nodules, isolated remnants, or associated cavities were rarely
observed. Furthermore, their opal-A spheres (also <10 nm to 50 nm in
diameter) tended to be more spherical in shape.
 Microfacies
3 sinter (Figure 3) is confined
to the southern portion of the study site, which slopes at ~10° E from the
horizontal. Although these deposits are usually submerged under flowing
thermal water (Figure 15), the water
level in this area is lower (~6-10 mm) than that flowing on flatter areas
(>10 mm) elsewhere at the site (Figure
4.1). Therefore, this microfacies is affected by changing water levels,
completely exposing the sinters to the air in times of lower water levels.
Water temperatures range from ~60-54°C (Figure
4.1).
Microfacies
3 sinter (Figure 3) is confined
to the southern portion of the study site, which slopes at ~10° E from the
horizontal. Although these deposits are usually submerged under flowing
thermal water (Figure 15), the water
level in this area is lower (~6-10 mm) than that flowing on flatter areas
(>10 mm) elsewhere at the site (Figure
4.1). Therefore, this microfacies is affected by changing water levels,
completely exposing the sinters to the air in times of lower water levels.
Water temperatures range from ~60-54°C (Figure
4.1).
Sinter from this environment is flat and
parallel-laminated (Figure 15.1).
Surfaces are irregular and even rippled in places, which appears to be due
to the patchy nature of ongoing silica deposition (Figure
15.2). During lower water levels, small, isolated puddles of thermal
water occur in the irregular surface crevices of the sinters. Continuous
patchy silica deposition on these sinters results in silica rims (Figure
15.3) that eventually grow into cup-like deposits like those of
Microfacies 2 (Figure 15.4).
Indeed, pumice clasts that rest partially submerged in these waters and on
top of the planar sinter deposits act as substrates for Microfacies 2
sinters (Figure 15.1). Due to the
thinness of Microfacies 3 and 4 sinter (~<3 mm and 2 mm,
respectively), no thin sections were made of these two deposit types.
Because of difficulties in permanently placing glass slides horizontally,
silica accretion rate measurements were also not conducted for this
microfacies. Any silica that deposits on vertically oriented slides would
have represented Microfacies 2 conditions.
 SEM
revealed that the upper surfaces of Microfacies 3 sinters are
colonised by bacilli (1-8 µm long) (Figure
16.1). Silicification of these microbes was patchy, with nanospheres
(<100 nm) of silica precipitating onto the uppermost portions of cells,
whereas those beneath were unsilicified (Figure
16.2). Vertical sections of the sinters reveal silica with a
predominantly massive, vitreous texture (Figure
16.3). Cavities, cemented diatoms, and spherical cells (2-10 µm in
diameter), resembling those from the green living mats, are the only
discernable features in these sections (Figure
16.3-16.4).
SEM
revealed that the upper surfaces of Microfacies 3 sinters are
colonised by bacilli (1-8 µm long) (Figure
16.1). Silicification of these microbes was patchy, with nanospheres
(<100 nm) of silica precipitating onto the uppermost portions of cells,
whereas those beneath were unsilicified (Figure
16.2). Vertical sections of the sinters reveal silica with a
predominantly massive, vitreous texture (Figure
16.3). Cavities, cemented diatoms, and spherical cells (2-10 µm in
diameter), resembling those from the green living mats, are the only
discernable features in these sections (Figure
16.3-16.4).
On vertical portions of the sinters that
have subaerial rims (Figure 15.3-15.4),
diatoms clustered together in large groups with the tips of their tests
aligned perpendicular to the approximate air-water interface (Figure
16.5). These clusters also became progressively silicified and
incorporated into the deposit (Figure
16.6).
 Sinters
from Microfacies 4 (Figure 3)
are characterised by thin (~2 mm thick), orange, cup-like rims formed on
small (<2 cm in diameter) pumice clasts that rest upon moist sandy
substrates well above the outflow channels (Figure
17.1-17.3). The deposits can reach heights of 1-6 mm, with rare or
absent spicular textures. Vertical sections through the rims reveal thin
internal laminae (~0.5 mm thick) that are convex rather than horizontal (Figure
17.4). The outer sides of these sinters are typically covered with green
algal mats. Rarely, anastomosing ridges occur on the inner sides of the
sinter rims (Figure 17.4).
Sinters
from Microfacies 4 (Figure 3)
are characterised by thin (~2 mm thick), orange, cup-like rims formed on
small (<2 cm in diameter) pumice clasts that rest upon moist sandy
substrates well above the outflow channels (Figure
17.1-17.3). The deposits can reach heights of 1-6 mm, with rare or
absent spicular textures. Vertical sections through the rims reveal thin
internal laminae (~0.5 mm thick) that are convex rather than horizontal (Figure
17.4). The outer sides of these sinters are typically covered with green
algal mats. Rarely, anastomosing ridges occur on the inner sides of the
sinter rims (Figure 17.4).
Thermal water was not observed to wash,
splash, or spray onto these deposits. Small holes dug into the sandy
substrates revealed warm thermal pore water seeping upward through the sands
(Figure 17.3). This water may derive
from surrounding thermal discharges nearby, or from small unidentified vents
underneath the sandy alluvium. The temperature of the seeping water was
recorded to range from 45-67°C (Figure
4.1). Silica accretion rates in this microfacies averaged only 9.5 x 10-3
g over 67 days (Figure 4).
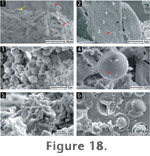 Sinter
surfaces are covered by biofilms of diatoms (Figure
18.1-18.2), spherical microorganisms (2-6 µm in diameter) (Figure
18.3-18.4), and bacilli (1-2.3 µm long) (Figure
18.5). It is likely that the spherical cells are also algae, belonging
to the green mats that cover the sinter rims on the margins of the deposits.
Microbial silicification is likewise patchy, with some microbes completely
covered in silica spheres, while others nearby are unsilicified. The bacilli
also had the tendency to form vertically upright microcolonies, but were
rarely observed to be silicified (Figure
18.5).
Sinter
surfaces are covered by biofilms of diatoms (Figure
18.1-18.2), spherical microorganisms (2-6 µm in diameter) (Figure
18.3-18.4), and bacilli (1-2.3 µm long) (Figure
18.5). It is likely that the spherical cells are also algae, belonging
to the green mats that cover the sinter rims on the margins of the deposits.
Microbial silicification is likewise patchy, with some microbes completely
covered in silica spheres, while others nearby are unsilicified. The bacilli
also had the tendency to form vertically upright microcolonies, but were
rarely observed to be silicified (Figure
18.5).
SEM showed that vertical sections of these
sinters comprise a massive, vitreous texture with no visible laminae.
Silicified spherical cells (2-6 µm in diameter), often at the endospore
stage, were commonly incorporated into the sinter (Figure
18.6). However, these organisms do not form layers as in Microfacies
2, but are scattered across the sinter.