|
|
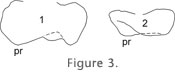
METHODOLOGICAL ISSUESProfile VisibilityOur curves were drawn over protocone-paracone and protocone-metacone in the upper molars, and protoconid-metaconid and hypoconid-entoconid in the lowers. We had first considered using the anterior or posterior wall of the specimen, anteroloph(id) and posteroloph(id) respectively, but that profile was quite variable within a population.
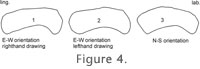
In some cases, we substituted a profile of the posterior border of the tooth (the posterolophid) because it was close to the profile we needed (e.g., the lower molars of B. bravoi). In other cases (E. quercinus, B. bruijni) the posterior profile was not visible in posterior view, but it could be drawn reliably in anterior view. In some species (Glamys) there was no connection between hypoconid and entoconid, and we constructed a composite curve of hypoconid, posterolophid, and entoconid for m1inf_post and m2inf_post. ParallaxIn the following paragraphs we use North-South and East-West to describe the axes of a microscope field as if it were a compass. When we began drawing the curves, we oriented the specimens E-W, with the occlusal surface facing N. When we tested the reliability of curves by having various people draw the same specimen, we were confronted with a serious problem: in some cases there was no similarity whatsoever between a curve drawn by a right-handed person using the camera lucida on the right side of the microscope and a curve drawn by a left-handed person working on the opposite side. Exactly the same problem occurred when the same person drew the same specimen with the occlusal surface facing N then with it facing S.
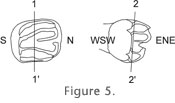
A solution might be to use a non-stereoscopic microscope, but that is not a common instrument among students of fossil rodent teeth, and its relatively shallow depth of field would cause other problems. We solved this problem by orienting the specimen N-S, with the occlusal surface facing to the side where the camera lucida is mounted. The depth of the concavity (h) is now oriented E-W, and if there is a distortion it affects the small value h and not the large value of the curve basis (b), and, if h is not a horizontal line, its inclination is always less than that of b. Furthermore, by facing the occlusal always to the side of the camera lucida, the distortion, if present, acts always in the same direction, regardless whether one draws left-handed or right-handed. It is evident that such a standardized orientation is fundamental for obtaining mutually comparable results. Vertical Orientation
Evidently the orientation of the specimen is important. This became clear while digitizing the eight anterior curves of B. micio published by Vianey-Liaud (1994; fig. 16) (= M-numbers in Table 6), and comparing them with our curves from specimens of the same species in the RGM collection. The results are given in Table 6. Among the M-numbers, three specimens are very concave, and five are much less so. Such a difference may well be due to the degree of wear, but the extremely high values of the three concave profiles could not be replicated by us using unworn specimens of the RGM collection. As a further experiment, we drew one specimen vertically, which yielded lower concavity values than the three very strongly concave profiles, and then drew the same specimen in an inclined position of about 65°, which yields much higher concavity values (see Table 6; values for h/r 0.24 versus 0.34). Apparently a false impression of concavity may be given when the orientation does not coincide strictly with the optical axis of the microscope tube. This inclined orientation may well explain the high values for the three mentioned specimens. In fact, in the drawings by Vianey-Liaud (1994) one may observe the anteroloph drawn below the deepest point of the protoloph. In the RGM specimen in the (sub)vertical position, part of the profile of the protoloph has to be estimated because it is hidden by the anteroloph. So, probably, the specimens drawn by Vianey-Liaud were oriented obliquely to make the protoloph visible. Alternative Orientation
The problems of parallax distortion and visibility are inherent to the kind of microscope used. We considered orienting the specimens horizontally and using a microscope that allows vertical measurements. However, we were looking for a method that would be applicable for any researcher using the normally available stereoscopic microscope. A method using a confocal microscope, for example, would not be available to students who don't have access to such a special instrument. It cannot be denied, however, that the precision of a more expensive alternative would better control the problem of parallax than the method we present here. Reliability and RepeatabilityThe orientation of the specimens inevitably implies a certain amount of error; therefore, a specimen of M2 of O. libanicus from MLB8 was drawn several times by various people, each time after moving and reorienting the specimen. The results are given in Table 7, and give a good idea of the reliability of the method. The last row of the table gives the variation of each parameter as a percentage of the midpoint of the range. The error of orienting and drawing the specimen causes a variation of less then 15% on either side of the midpoint. The profiles of M. misonnei, drawn by Vianey-Liaud (1994), were analyzed and compared with the profiles of several specimens from the RGM collection drawn by the present authors. The results are quite comparable. As a further test, the anterior profiles of eight specimens of M2 of B. itardiensis from OLA4A were drawn and vectorized, in order to estimate the variation within a population. Table 8 represents the values obtained for each specimen and the mean and standard deviation of the various parameters. We visually inspected additional specimens to determine whether our samples were adequate to characterize the typical concavity within a species. A total of 26 M1 of O. libanicus from MLB8 were inspected without drawing them; 12 specimens were unworn and showed no obvious differences in concavity; among the remaining 14 specimens not a single one made us suspect an appreciably different degree of concavity. In order to test how well the circle coincides with the profile, the value of dev as defined in the Methods section, was divided by the number of vertices (v) that constitute the polygon. The deviation from the circle is on average slightly higher in the lower molars than in the upper ones (0.15 vs. 0.10), but the difference is not significant at a 5% confidence limit. Also, for each profile a T-test was executed on the deviations from the circle. Among 510 profiles only 18 deviate significantly from a circle (at a 5% confidence level). These 18 profiles were equally distributed among lower and upper molars and among weakly and strongly concave profiles. Though this method makes it possible for the first time to quantify concavity objectively, we note that orientation of the specimens is inevitably a subjective factor that may have considerable influence on the results. A partial solution to the problem of orientation would be to draw each specimen several times, and use the mean values calculated from these drawings. Degree of WearCrown wear obviously influences observed concavity. Whenever possible we used unworn or minimally worn specimens, but such ideal specimens are not always available. Significantly worn specimens were always discarded, but in intermediate cases we found that it was possible to plausibly reconstruct the missing part of the worn cusps. Normally wear affects the delimiting cusps first, and one may expect a gradually decreasing concavity from unworn to very worn specimens. The case of B. concavidens seems to indicate that there may be an increase in concavity during the first stages of wear (see the discussion of Bransatoglis in the Results section). SkewnessThough not a measure of concavity, we also measured the skewness of the profiles, as defined by b1 and b2; these values are identical in symmetric profile. In a very asymmetric curve b1 and b2 are substantially different. The skewness of the curve was measured as the percentage of b that is occupied by b1, or, in other words, 100 x b1/b. Values around 50 indicate symmetric curves, low values mean that the deepest point of the concavity is lingual of the molar axis in the upper molars (labial of the axis in the lower molars); high values indicate the opposite. When we started our investigation we decided to measure skewness, in order to find out if it played a role in concavity, because both skewness and concavity may play a role in different ways of occlusion. There is no apparent regularity in the distribution of the skewness. One would expect that there were a certain relation between the skewness value for the posterior profile of M1 and the anterior profile of M2, but that is not the case. Nor is there an evident pattern of distribution between the skewness of upper and lower profiles. Further study will have to show whether measuring skewness has some significance. |
|