| |
DISCUSSION
The family Synchytriaceae (Chytridiales) includes over 200 species, most of which are contained within the genus Synchytrium, that are obligate parasites on flowering and non-flowering plants, as well as on mosses and algae (Karling 1964;
Lutzoni et al. 2004). The family is also characterized by a colonialist developmental type, and by the formation of thin-walled and/or thick-walled sporangia. Some species are recognized by the fusion of isogamous gametes. Within Synchytriaceae, several other fungal characteristics, such as type of life cycle, size, shape, cytology, and structure of zoospores, sporangia, and resting sporangia are sometimes useful to discern Synchytrium subgenera and species (Karling 1964). In addition, host range, cellular host reaction, and environmental conditions of the site of occurrence have been used in their identification and classification (Karling 1964). The complete life cycle of extant Synchytrium is only known in a few cases (Curtis 1921;
Kusano 1930;
Karling 1964), sometimes precluding a narrow morphological comparison and subsequent separation of species. In spite of this, the presence or absence of sporangial sori, resting sporangia, development of the former from the initial thalli or from prosori, and the germination method of resting sporangia (as a prosorus or as a sporangium), have aided in separating Synchytrium into six subgenera (Karling 1964). Whether the initial cell or vegetative thallus gives rise to a prosorus, a sorus, or a resting sporangium, Synchytrium species which lack either resting sporangia or sporangial sori are termed short life cycle, whereas when these structures are produced the species are considered long life cycle (Karling 1964).
Extant fungi or fungus-like endoparasites produce zoospores as an alternative means of dispersion only when enough moisture characterizes the environment, otherwise species adapted to dry environments germinate directly through a germ tube without producing zoospores unless conditions become moist enough for zoospore dispersal (see
Webster and Weber 2007 and references therein). Accordingly, the presence of a large number of zoospores in the Antarctic Late Permian assemblage indicates a moist environment, at least one with enough moisture locally so that dispersion through means of swimming zoospores was possible. Additionally, the abundance of fungi (including rot fungi, a Glomus-like fungus, and other assignable to basiodiomycetes)
(Stubblefield and Taylor 1986;
Visscher et al. 1996;
Diéguez and López-Gómez 2005;
García Massini 2007) in this Late Permian assemblage may be a reflection of the abundance of resources (i.e., abundant organic debris for fungal saprotrophs),
something that has been suggested for other assemblages of comparable age
worldwide.
Widespread fungal infections in fossil forest stands in environments subjected to recurrent ecological disturbance have been suggested to debilitate plants, possibly making them increasingly more susceptible to pathogenic fungi (opportunistic pathogens) (Creber and Ash 1990;
Falcon-Lang and Cantrill 2001). Regarding this, it has been indicated that the amount of resources used by extant pathogenic fungi to disperse their propagules decreases with increasingly higher density and geographic proximity of their host(s) (Damgaard 1999). Late Permian environments worldwide were subjected to various types of disturbance that, in turn, have been indicated to be singly or in combination the cause(s) of the massive extinction that affected the biota at the Permian-Triassic boundary (e.g., see
Hsü and McKenzie 1990;
Knoll et al. 1996;
Looy et al. 1999;
Retallack et al. 2005). The Late Permian assemblage from which the fungus described here comes was characterized by a homogeneous pteridosperm flora of the Glossopteris type that lived in an environment subjected to volcanic activity, and this combination of factors can be suggested to have bolstered the occurrence of fungi, including pathogenic individuals (Pigg 1990;
Taylor et al. 1992;
Pigg and Taylor 1993).
Based on the endobiotic parasitic habit, the production of thin- and thick-walled sporangia and zoospores in different states of development the fossil described here is most similar to extant members of Synchytriaceae, particularly to Synchytrium, and are therefore assigned to a new species, termed permicus. Additionally, a life cycle can be hypothesized for the fossil endoparasite and this appears most similar to the long life cycle type (Karling 1964). However, differences, such as the occurrence of a single sporangium per host cell in the fossil, as opposed to a colonialist habit in modern taxa where a cluster of sporangia is present in a single host cell, are noted. Other characteristics of the Permian fungus are coincident with those displayed by members of Synchytrium, particularly Mesochytrium, though these are too variable in the fossil, and rarely diagnostic in modern species (Karling 1964). For example, thick-walled sporangia may be ellipsoid, ovoid, or pyriform, and possess a wall that is of variable thickness and externally ornamented with pits or ridges; thin-walled sporangia may be nearly spherical and formed by numerous small polygonal segments or ellipsoid and built of several transversely arranged segments. This variability is included within the range of morphological variation present in extant Synchytrium species (Figure 2.1, 2.2., 2.3, 2.4,
Figure 3.5, 3.6, 3.7, 3.8). Also, the zoospores (Figure 1.3, 1.4, 1.5, 1.6, 1.7, 1.8) have a wide shape and size range. These features are present in members of Synchytrium, although they may be highly dependent on environmental conditions, density of parasites in the host, and degree of development of the infection, and alone, therefore, may not be diagnostically reliable (Karling 1964).
Additional information can be inferred from the many fossils preserved and this supports affinities with Synchytrium. Many zoospores are preserved within the Permian peat in many different plant fragments showing varying degrees of decay, and these could also have been produced by a different endoparasitic fungus, including oomycetes and plasmodiophoromycetes. However, diagnostic structures of these two latter groups, such as oogonia or antheridia for oomycetes, or sporosori with a determinate number of resting spores, each containing a single biflagellate zoospore, for plasmodiophoromycetes, are missing and so their presence might only be hypothesized. Meanwhile, uniflagellate zoospores, thin- and thick-walled sporangia containing zoospores, and zoospore morphs in different stages of fusion are diagnostic for Synchytrium (Karling 1964;
Barr 2001;
Dick 2001;
Brasselton 2001; and see
Webster and Weber 2007 for additional references). Other characteristics of these structures (e.g., shape, size, and thickness of their wall components) are comparable among these three groups and do not provide evidence for distinguishing either. Consequently, morphology alone is not the most accurate means to identify modern or fossil members of Synchytrium; for this reason, if available, other criteria should be used. In Synchytrium permicus, the large number of specimens in different stages of development provides the opportunity to speculate on the biology of this fossil fungal endoparasite with a long life cycle type.
Hypothetical Life Cycle
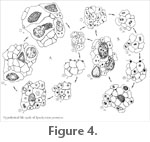 The life cycle of Synchytrium permicus
as an endobiotic fossil parasitic fungus with a holocarpic and monocentric thallus can be postulated based on the range of development stages of specimens associated with parasitized silicified plant remains from the Permian of Skaar Ridge (Figure 1.1). The life cycle would have begun with the liberation of zoospores from thin-walled sporangia, some of which acted as isogamous gametes (haploid) that fused sexually forming thick-walled sporangia (diploid) that produced additional zoospores (Figure 4). The life cycle of Synchytrium permicus
as an endobiotic fossil parasitic fungus with a holocarpic and monocentric thallus can be postulated based on the range of development stages of specimens associated with parasitized silicified plant remains from the Permian of Skaar Ridge (Figure 1.1). The life cycle would have begun with the liberation of zoospores from thin-walled sporangia, some of which acted as isogamous gametes (haploid) that fused sexually forming thick-walled sporangia (diploid) that produced additional zoospores (Figure 4).
Liberation and encystment of zoospores. The life cycle begins with the liberation of zoospores (haploid) from the thin-walled sporangia. Two different patterns of liberation are suggested; one in which zoospores are separated and individually discernible from each other, and one in which they form a more or less cohesive group resembling a stream of zoospores (Figure 1.2,
Figure 2.5,
Figure 4.1). An embedding gelatinous substance during liberation of zoospores from sporangia occurs in fungi such as
Allomyces and Entophlyctis, and might be characteristic of advanced species (Powell 1976;
Duff and Youatt 1977;
Barr 1978). It has been suggested that whether released zoospores in Synchytriaceae appear individually or as a compact mass is a function of the state of maturity (Curtis 1921;
Karling 1964).
Wide variation in zoospores size and shape occurs in S. permicus (Figure 1.3), and similar variation occurs in modern Synchytriaceae (Powell 1993;
Karling 1964). Some of the larger fossil zoospores may represent encysted zoospores or developing sporangia (i.e.,
Figure 1.8). The centrally located opaque contents in the fossil zoospores (Figure 1.4) resemble the internal contents seen in extant taxa (Lange and Olson 1978;
Dewel et al. 1985).
After adhering to the host cell surface (Figure 4.2, 4.3), the zoospores of S. permicus
appear to have formed a hemispherical structure firmly attached to the host cell (Figure 1.6,
Figure 4.3). An opaque, thick, short knob at the base of some zoospores may represent the remnant of the flagellum once encystment started (Figure 1.4,
Figure 4.3), as in modern Synchytriaceae (Karling 1964). One or more opaque threads extending from the central body towards the periphery of the fossil zoospores (Figure 1.5,
Figure 4.3) are interpreted as remnants of the flagellar apparatus; or this may represent the transference of the zoospore contents into the host cell. Similar threads arise from the nucleus and represent organelles disaggregated during encystment and penetration in modern Synchytrium fulgens and Synchytrium endobioticum (Curtis 1921;
Kusano 1930). A few empty zoospores suggest that only the organelles and nucleus were transferred into the host cell (Figure 1.3,
Figure 4.4). However, since other zoospores occurred intact within host cells it is not possible to determine whether one of the two mechanisms prevailed (Figure 4.4). Examples of both transference of zoospore contents leaving an empty sheath or membrane outside the host cell, as in S. australe, and ingression of the entire zoospore into the host cell, as in S. endobioticum, occur in modern Synchytriaceae (Curtis 1921;
Karling 1955).
Behavior of zoospores inside the host. Large zoospores, probably encysted individuals, or developing sporangia indicate that zoospores and their contents enlarged while maintaining their size ratio and
spherical shape (Figure 1.8,
Figure 4.4, 4.5). A few enlarged zoospores appear elongated, which may represent movement across the host cell (Figure 1.7,
Figure 4.5), as in some extant Synchytriaceae before encystment (Karling 1964). Large encysted individuals have variable amounts of opaque contents and an irregular scar-like aperture on the surface (Figure 1.8, 1.9,
Figure 4.5), suggesting that the encysted zoospore or developing sporangium contents may have been discharged into the host cell. The presence of host cells filled with two similarly sized diffuse bodies, one containing opaque inclusions, also suggests zoospore content migration (Figure 1.10,
Figure 4.5). This resembles some stages of migration of the contents of encysted zoospores (i.e., prosori) into host cells in several extant members of the Synchytriaceae (Karling 1964).
Formation of thin-walled sporangia. Several sporangia of various sizes suggest that in the next stage the encysted zoospore or developing sporangium started a segmentation period that assumed two different patterns (Figure 4.6). In one pattern the lumen appears to have cleaved forming numerous polygonal segments of approximately equal size (Figure 2.1,
Figure 4.6). Each of these segments, some of which still have part of their wall preserved, is interpreted as a zoospore primordium (haploid). In this instance, the host cell and the sporangium are nearly spherical
(Figure 2.1,
Figure 4.6). Liberation of zoospores (haploid) from this type of sporangium probably
occurred after the walls of both host cell and its infecting sporangium swelled
and burst (Figure 2.1,
Figure 4.1). In the other pattern, it appears that the lumen of the sporangium was invaginated to form several transversely arranged globose segments of unequal size that partially divided it (Figure 2.2,
Figure 4.6). Other sporangia have a fully divided lumen formed of smaller globose segments that suggest that these further cleaved (Figure 2.3,
Figure 4.1).
What are interpreted as mature sporangia are ellipsoid to broadly ellipsoid individuals
(Figure 2.3, 2.4,
Figure 4.1) with an acute papilla-like body projection (Figure 2.4,
Figure 4.1). This papilla might have detached from the wall leaving an opening through which zoospores (haploid) were liberated. A few ruptured sporangia suggest that zoospores were liberated by disruption of the sporangial wall (Figure 2.5,
Figure 4.1). Once zoospores (haploid) were liberated, they might have either immediately encysted to form thin-walled sporangia or fused and then encysted to form thick-walled sporangia (Figure 4.2, 4.3, 4.7).
In almost all sporangia examined, what appears to be remnants of the sporangial wall or sporangial lumen or cytoplasm remained after zoospore liberation (Figure 2.1, 2.5). Similar structures, which represent irregularly cleaved anucleated segments of cytoplasm, occur in Olpidium brasssicae (Temmink and Campbell 1968). Polygonal segments in some of the fossil sporangia resemble segments in modern chytridiomycetes that result from the initial cleavage of the sporangium (Karling 1977). The sometimes globose to irregularly shaped segments seen in the fossil usually are not seen in modern chytridiomycetes; however, in several members of Synchytriaceae (i.e., S. taraxaci, S. endobioticum) the cleavage planes are laid down in a non-synchronous manner, producing irregular segments (De Bary and Woronin 1863;
Curtis 1921). Whether rounded or polygonal, several authors have suggested that the shape of segments is largely dependent on the size of the host cell (Temmink and Campbell 1968).
Only one sporangium per host cell was detected in the fossil. Such monosporangiate reproductive structures are rather unusual in the Synchytriaceae, but have been seen in some species (Curtis 1921). Moreover, a developing sorus in Sorochytrium milnesiophthora may result in a monosporangiate reproductive structure if the host dies prematurely (Dewel et al. 1985). In other chytridiomycetes the shape remains constant until partitioning of the sporangia into individuals begins (Karling 1964,
1977). Formation of a discharge papilla and its detachment from the sporangial wall as well as rupture of the sporangium for zoospore liberation is common in chytridiomycetes (Barr 2001). The papilla-like body projection in the sporangial wall of the fossil is nearly identical to the discharge papilla formed as an extension of the enveloping membrane or sporangial wall in plant parasites such as O. brassicae, S. endobioticum, S. taraxaci (De Bary and Woronin 1863;
Curtis 1921).
Fusion of zoospores and zygote maturation. Several zoospores (haploid) closely appressed to each other in S. permicus may be an early stage of sexual development seen in modern Synchytrium species (Karling 1964). These pairs of similarly sized zoospores appear close to each other, whereas other pairs appear attached, both with individual margins distinguishable (Figure 2.6, 2.7,
Figure 4.7). Other pairs of elongated, equally sized zoospores attached to each other, but with a common margin, may represent a further step in the formation of the zygote (diploid) (Figure 2.8,
Figure 4.8). In these individuals, the central opaque contents of each zoospore individually adopt an ellipsoid shape, and subsequently fuse to form a single body (Figure 2.8,
Figure 4.8, 4.9). Accompanying these pairs of zoospores are larger cells with an approximately circular opaque inclusion, which might represent zygotes (diploid) after fusion is complete (Figure 2.8,
Figure 4.10). The fossil zoospores occur in plant tissue, on which the zygotes would have encysted.
Fossil zoospores interpreted as fusing gametes (haploid) are nearly identical to several stages of zygote formation in species such as S. australe, S. fulgens,
and S. endobioticum (Curtis 1921;
Kusano 1930;
Karling 1955). In the modern species the fused nuclei become circular with a flattened contact surface, which first appears opaque and later translucent. In contrast, the opaque bodies in the fossil zoospores become ellipsoid during fusion (Figure 2.8,
Figure 4.8, 4.9). Some of the fossil zygotes exhibit what is interpreted as a single thin flagellum or its remnants (Figure 2.9,
Figure 4.10). This may represent fusion between a motile and a non-motile zoospore (haploid), or may be the result of poor preservation. A few large zoospores appear to be zygotes (diploid) with opaque inclusions of dense material in the cytoplasm (Figure 3.1, 3.2,
Figure 4.10, 4.11). These resemble different stages of nucleus rearrangement during zygote maturation in Synchytriaceae (Karling 1964).
Formation and maturation of thick-walled sporangia. The host tissue commonly appears infected with several large zoospores and thick-walled sporangia in different states of development (Figure 1.1,
Figure 4.11). Some of the larger zoospores may have been zygotes (diploid) that developed into thick-walled sporangia. As a result of the infection, a more or less central agglomeration of sporangia relative to the location of zygotes is observed (Figure 1.1,
Figure 4.11). Host cells containing the larger sporangia appear hypertrophied (Figure 2.2,
Figure 3.6), whereas those containing the smaller individuals sometimes appear distorted (Figure 3.4). Several modern fungal plant parasites produce various host reactions including hyperplasia and hypertrophy (Karling 1964;
Barr 1992,
2001;
Powell 1993).
The thick-walled fossil sporangia vary widely in size and shape (Figure 3.2, 3.3, 3.4, 3.5, 3.6). Such variation occurs in modern Synchytriaceae and depends on the size and type of host cell, as well as on the number of sporangia in the host cell (Karling 1964). The sporangia are similar to those of other plant fungal or fungal-like endoparasites, such as Polymyxa, Physoderma, Olpidium, and Synchytrium (Karling 1964;
Temmink and Campbell 1968;
Sparrow 1975). In particular, ridges ornamenting the perimeter of the sporangial wall in the larger sporangia, and a warty to pitted surface in the smaller ones are seen in S. endobioticum and S. fulgens, respectively (Curtis 1921;
Kusano 1930; also see
Karling 1964 and
Webster and Weber 2007) (Figure 3.4, 3.6). In modern Synchytriaceae, the ornamentation and apparent wall layers are, in part, a consequence of accumulation of dead material from the host cell (Karling 1964).
Several distorted, thick-walled sporangia suggest irregular rupture of the sporangial wall as means of zoospore liberation (Figure 3.7,
Figure 4.12). Liberation also may have occurred through one small slit located at the end of the thick-walled sporangium (Figure 3.5,
Figure 4.12). A long maturation period during which sporangia and zoospores develop and grow in size before they are liberated through a slit or a complete rupture of the sporangial wall was described in Synchytriaceae (Karling 1964). After fossil zoospores were released from the sporangia they may have encysted creating new thin-walled sporangia, or perhaps fused, and then infected a new host cell to create additional thick-walled sporangia (Figure 4.2, 4.7).
|