|
|
|
DISCUSSION AND FUTURE WORKThis model revealed that in order for the primary movements to occur, as described in the original pleurokinetic hypothesis (Weishampel 1984) (Figure 3), extensive secondary movements must also occur. Moreover, some of the secondary movements observed involved large (> 1 cm) separation of the elements, including movements between the jugal and quadratojugal, pterygoid and palatine, and pterygoid and quadrate (Figure 10). Importantly, these movements arose with only small amount of abduction of the maxillae and quadrate (e.g., 3º, Table 1), which were much less than those in the original Corythosaurus model. Presumably, the pleurokinetic model, as originally proposed by Weishampel (1984) would have yielded secondary movements much greater than those observed in the Edmontosaurus model presented here. Both the original model and this one did not incorporate soft tissues such as ligaments, synovial capsules, and musculature though their inclusion would greatly enhance functional resolution and allow for more rigorous hypothesis testing. Ligaments, including those within sutures and larger ones, such as the quadrate-articular ligament if present, may have constrained some, if not much of the movement in the facial skeleton this model illustrated. In particular, it seems initially doubtful that such separation of cranial elements is likely to have taken place in the living animal considering that sutural ligaments are comprised of collagen, which yields at only 4 to 5% strains (measured in tendon, Wainwright et al. 1976). Additionally there is a significant amount of bony overlap between many of these articulations, suggesting that these contacts promoted stability rather than mobility. Similarly, synovial capsules were likely present, minimally at the otic (quadrate-squamosal), basal (pterygoid-basisphenoid), and jaw joints. These capsules would have potentially constrained movement expressed in the model. The significance of these factors would depend on the functional properties assigned to them at different time stages of the model, requiring multiple assumptions.
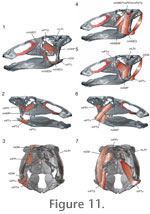
In addition, many of the aforementioned jaw muscles attach across a number of the elements and joints that were implicated in secondary movements. The lateral surfaces of the palatine, pterygoid, and quadrate are a likely attachments for M. pterygoideus dorsalis and M. adductor mandibulae posterior, whereas the medial surfaces of these elements were probably attachment regions for M. pterygoideus ventralis and M. protractor pterygoideus (Figure 11). This pattern of muscle attachment suggests that the palatine, pterygoid, and quadrate formed a functional unit, providing a supposedly stable, immobile attachment for jaw musculature. Yet the model exhibited extensive sliding between the quadrate and pterygoid during chewing challenging functional hypotheses regarding how this part of the skull may have simultaneously served to produce and resist chewing forces. Therefore, jaw musculature, if modeled, would not only provide the power to the modeled feeding apparatus, enabling analyses of acceleration and bite force moments, but would also induce stresses across sutures and on the bones of attachment themselves, enabling analysis of deformation (e.g., via finite element modeling), and hypotheses of mobility (versus stability) to be tested. Full consideration of the soft tissue systems may in the end reveal that maxillary abduction is not a viable mechanism for producing transverse chewing movements. With this in mind, additional insight into the potential mobility of the hadrosaurid quadrate may be derived from study of other hadrosaurid taxa. Indeed, a review of the cranial joints of Brachylophosaurus, another hadrosaurine dinosaur, found that the morphology of the pleurokinetic hinge regions (e.g., quadrate-squamosal joint) would have prohibited maxillary abduction (Cuthbertson 2006). Therefore cranial kinesis may be variable among hadrosaurs, or perhaps the hadrosaurid skull was less mobile than originally assumed. However, part of the difficulty in analyzing ornithopod chewing is that the proposed transverse abduction of the maxillae is rare, if not unique among amniotes, so identifying appropriate analogs is difficult. In ongoing work we plan to further correct parts of the skull model, which may be still deformed, as suggested by the fact that during the powerstroke the teeth are not in precise occlusion (Figure 9). We will also investigate the role of soft tissues in constraining the model (Ostrom 1961, Holliday 2006). The system will be further constrained by incorporating dental wear data including the pattern of scratches (i.e., microwear striations) and orientation of the occlusal surface. If these wear features prove to be well-preserved they will provide an accurate map of the relative movements of the upper and lower tooth rows (e.g., Rybczynski and Reisz 2001). The inclusion of the additional functional parameters will allow future analyses to constrain the range of possible chewing movements (e.g., see Hutchinson and Gatesy 2006) to gauge how sensitive the model is to different variables. With these additional constraints in place, alternative hypotheses of feeding behavior can be more fully explored including simple quadrate streptostyly, akinesis, and mandibular propaliny or rotation. In conclusion, preliminary results suggest that the integration of three-dimensional scanning technology with animation offers enormous promise for investigating the functional morphology of feeding behavior in Edmontosaurus. Although the goal of the methods described here is to resolve the feeding kinematics, the approach lays the groundwork for investigating questions related to biomaterials and chewing forces across all vertebrates. Ultimately, animation and biomechanical methods could be applied comparatively to yield insight into the evolution of feeding systems within lineages and ecosystems. |
|