| |
Methods
Creation of three-dimensional conceptual models of trace fossils differs greatly
from the process of direct reconstruction of the three-dimensional morphology of
fossil material based on serial grinding and tomography. This paper aims to
produce a three-dimensional deterministic model of some phycosiphoniform burrows
from turbiditic siltstone of Cretaceous Rosario Formation and compare them to
the Phycosiphon model of
Bromley (1996). The approach used involves the
use of serial grinding and computed tomography as outlined below.
Serial Grinding
Three-dimensional geometry of the studied burrows was systematically exposed
through serial grinding of the hand specimen. This approach has been
successfully employed for three-dimensional imaging of body fossils (e.g.,
Baker
1978; Hammer 1999;
Sutton et al. 2001), ichnofabric (Wetzel and Uchman 1998,
2001) and trace fossils (Naruse and Nifuku 2008). Serial grinding allowed us to
obtain a sequence of regularly spaced images of the resultant vertical cross
sections. The photographic dataset thus created is the basis for subsequent
computer-based three-dimensional reconstructions.
To aid in creating parallel regularly spaced cross sections, the irregularly
shaped sample of turbiditic siltstone was placed in a tight fitting box and set
in plaster of Paris. When the plaster was set, and regular 0.5 mm increments
inscribed on the outer surface of the rectangular block, it was then ready to be
serially ground. The regular outline of the block was essential to create
reference points, for alignment of the photographic images to be used in digital
analysis. The 0.5 mm spacing of images was chosen to capture a sufficiently
large number of data-points to allow gridding of surfaces and reconstruction of
the burrows. A total of 60 images were acquired through a 29.5 mm thick slab of
the sample. The consecutive series of photographs were taken from parallel
surfaces with a digital camera, which was stationed an identical distance above
the sample surface, under the same lighting and zoom conditions for every
surface. The camera was attached to a photographic stand with height controlling
screw feed.
Ichnofabrics have not generally been studied using a serial grinding approach.
In contrast to body fossil material, trace fossil fabrics are commonly complex,
tortuous, and without sharply defined limits (both morphologically and
mineralogically). A particular problem is that burrows may branch and
inter-penetrate, making closely spaced slicing essential, and poses particular
challenges in image processing (discussed below). The size of the block studied
is larger than has typically been studied by palaeontologists, but did not pose
any particular methodological problems.
Image Processing
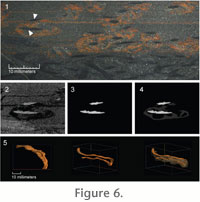 The set of sequential slice images acquired through the serial grinding
technique was processed to select the regions to be studied. The
phycosiphoniform burrows studied include a dark mud core and a halo of coarser
sediment, which in the present material is accentuated through the presence of
pyrite (Figure 1.1,
Figure 6.1). To obtain adequate contrast, the images were made into
gray scales (Figure 6.2). All images were put into a single Photoshop document
in consecutive order. Discrete burrow cores were chosen as the objects for
tracing the location of the chosen burrow. The burrow core was tracked through
each consecutive image and manually selected using layer masking to hide all
other burrow cores and halo that might confuse the reconstruction of the chosen
burrow (Figure 6.3). A masking layer was used to allow retention of the
original, gray scaled images, including location of adjacent burrows, should it
become subsequently desirable to study adjacent burrows (Figure 6.4). The
layered Photoshop document was then cropped to the smallest size that
encompassed the isolated burrow core. Each layer, representing the equidistant
ground surfaces, was saved as a JPEG image in the same directory with a numeric
name that indicates its position in the sequence. This set of image-processed
two-dimensional binary images was used for the subsequent three-dimensional
reconstruction. The set of sequential slice images acquired through the serial grinding
technique was processed to select the regions to be studied. The
phycosiphoniform burrows studied include a dark mud core and a halo of coarser
sediment, which in the present material is accentuated through the presence of
pyrite (Figure 1.1,
Figure 6.1). To obtain adequate contrast, the images were made into
gray scales (Figure 6.2). All images were put into a single Photoshop document
in consecutive order. Discrete burrow cores were chosen as the objects for
tracing the location of the chosen burrow. The burrow core was tracked through
each consecutive image and manually selected using layer masking to hide all
other burrow cores and halo that might confuse the reconstruction of the chosen
burrow (Figure 6.3). A masking layer was used to allow retention of the
original, gray scaled images, including location of adjacent burrows, should it
become subsequently desirable to study adjacent burrows (Figure 6.4). The
layered Photoshop document was then cropped to the smallest size that
encompassed the isolated burrow core. Each layer, representing the equidistant
ground surfaces, was saved as a JPEG image in the same directory with a numeric
name that indicates its position in the sequence. This set of image-processed
two-dimensional binary images was used for the subsequent three-dimensional
reconstruction.
Three-Dimensional Rendering
 The set of the binary images was imported to the commercial edition of VolView
2.0 software. Consecutive, gray scaled intersections of burrow core were
converted by the software to the volume shape that represents the
three-dimensional geometry of the examined phycosiphoniform burrow. Artificial
colors were attributed to the reconstructed burrow and to the halo in order to
aid illustration (Figure 6.5,
Figure 7,
Figure 8, and
Figure 9). Three-dimensional reconstruction of
the phycosiphoniform burrow from examined rock was additionally saved as a movie
file that shows the burrow rotating around the axis that is perpendicular to the
bedding plane (see attached animation files, Figure 7.7 and
Figure 9.4). The set of the binary images was imported to the commercial edition of VolView
2.0 software. Consecutive, gray scaled intersections of burrow core were
converted by the software to the volume shape that represents the
three-dimensional geometry of the examined phycosiphoniform burrow. Artificial
colors were attributed to the reconstructed burrow and to the halo in order to
aid illustration (Figure 6.5,
Figure 7,
Figure 8, and
Figure 9). Three-dimensional reconstruction of
the phycosiphoniform burrow from examined rock was additionally saved as a movie
file that shows the burrow rotating around the axis that is perpendicular to the
bedding plane (see attached animation files, Figure 7.7 and
Figure 9.4).
Three-Dimensional Morphology of the Rosario Formation Phycosiphoniform
Burrows
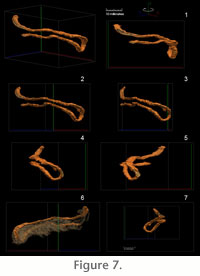
 By choosing a sparsely bioturbated portion of the ichnofabric, it was possible
to identify a single isolated burrow. The burrow consists of a single loop
shaped clay-filled tube that is identifiable in the series of ground vertical
cross sections. This isolated burrow was subjected to detailed three-dimensional
reconstruction of both the mud-filled burrow core (Figure 7.1-7.5) and the
burrow halo (Figure 7.6-7.7). The volume of rock subjected to three-dimensional
reconstruction, and containing the fossil burrow was 40.9 mm in length (X axis),
21.9 mm in height (Y axis) and 29.5 mm thick (Z axis) (Figure 7,
Figure 8, and
Figure 9). The two
limbs mud-filled burrow core that describe the shape of the lobe are parallel to
each other in vertical section and vary in diameter between 3 and 4 mm. Slight
thickening in tube width is noted in the distal portion of the loop that cannot
be attributed to compaction. Thickening of this part of the tube was described
as one of the diagnostic characteristics of Phycosiphon (Wetzel and
Bromley, 1994). The paired limbs of the examined form are not in the same
horizontal plane, and the terminal portion of the loop is at a steep angle to
the limbs. By choosing a sparsely bioturbated portion of the ichnofabric, it was possible
to identify a single isolated burrow. The burrow consists of a single loop
shaped clay-filled tube that is identifiable in the series of ground vertical
cross sections. This isolated burrow was subjected to detailed three-dimensional
reconstruction of both the mud-filled burrow core (Figure 7.1-7.5) and the
burrow halo (Figure 7.6-7.7). The volume of rock subjected to three-dimensional
reconstruction, and containing the fossil burrow was 40.9 mm in length (X axis),
21.9 mm in height (Y axis) and 29.5 mm thick (Z axis) (Figure 7,
Figure 8, and
Figure 9). The two
limbs mud-filled burrow core that describe the shape of the lobe are parallel to
each other in vertical section and vary in diameter between 3 and 4 mm. Slight
thickening in tube width is noted in the distal portion of the loop that cannot
be attributed to compaction. Thickening of this part of the tube was described
as one of the diagnostic characteristics of Phycosiphon (Wetzel and
Bromley, 1994). The paired limbs of the examined form are not in the same
horizontal plane, and the terminal portion of the loop is at a steep angle to
the limbs.
Nature of the Halo in the Rosario Phycosiphoniform Burrows
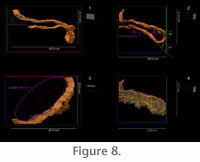
 Our 3D reconstruction of Phycosiphoniform burrows from the Rosario Formation,
Baja California, Mexico, demonstrates that the reworked silt-rich, clay-poor
material that forms the halo around the clay-filled burrow core is dominantly
present below the level of the clay-filled burrow (Figure 9). This feature is
also prevalent in most natural vertical cross sections studied in the field
(Figure 1.1). The halo is demonstrably meniscate, as determined from
cross-sectional views, but especially through three-dimensional reconstruction
(Figure 9). It is also noted that the burrow halos of adjacent burrow limbs are
closely juxtaposed with little if any undisturbed host sediment between them
(Figure 9). The halo around phycosiphoniform burrow cores has been described
from other occurrences (Wetzel and Wijayananda 1990;
Ekdale and Lewis 1991), but
has not previously been reconstructed in three dimensions. Our 3D reconstruction of Phycosiphoniform burrows from the Rosario Formation,
Baja California, Mexico, demonstrates that the reworked silt-rich, clay-poor
material that forms the halo around the clay-filled burrow core is dominantly
present below the level of the clay-filled burrow (Figure 9). This feature is
also prevalent in most natural vertical cross sections studied in the field
(Figure 1.1). The halo is demonstrably meniscate, as determined from
cross-sectional views, but especially through three-dimensional reconstruction
(Figure 9). It is also noted that the burrow halos of adjacent burrow limbs are
closely juxtaposed with little if any undisturbed host sediment between them
(Figure 9). The halo around phycosiphoniform burrow cores has been described
from other occurrences (Wetzel and Wijayananda 1990;
Ekdale and Lewis 1991), but
has not previously been reconstructed in three dimensions.
A similar halo associated with a phycosiphoniform burrow (attributed to
Anconichnus) was interpreted as an early diagenetic oxidation halo (Ekdale
and Lewis 1991). This feature was subsequently reinterpreted as being due to bioturbation, specifically the formation of spreiten in accord with newer
conceptual models (Wetzel and Bromley 1994;
Bromley 1996). Three-dimensional
reconstruction of the Rosario Formation phycosiphoniform fossil, with its
associated coarser-grained structure, demonstrates that the coarser-grained
material is indeed asymmetric and lies below the level of each of the two lobe
arms (Figure 9). This asymmetry is also visible from vertical surfaces prepared
in the laboratory and in natural outcrop (Figure 1). The burrow halo is
characteristically pyrite rich (Figure 1.2). Pyritization is interpreted to have
been caused by sulphate-reducing bacteria during early diagenesis. The marked
color contrast between the pyritized halo and clay-rich burrow cores relative to
the surrounding rock matrix allowed us to distinguish the three components of
the fabric for the purpose of image analysis.
The presence of the coarser-grained (silt-sized) material, not only between lobe
arms, but also external to the marginal tube (Figure 9) precludes the presence
of spreite and allows rejection of the possibility that the phycosiphoniform
trace fossil reconstructed herein is Phycosiphon. In the accepted
conceptual model of Bromley (1996;
Figure 3,
Figure 4, and
Figure 5), spreiten are predicted only
between arms of a single lobe and between marginal burrows. The behavioural
model proposed for Phycosiphon (Bromley 1996) precludes the possibility
of formation of the halo/spreiten below the level of a marginal tube that
borders the Phycosiphon structure. Spreiten are demonstrably not present
in our material from Rosario Formation. Instead, the phycosiphoniform cross
sections are inferred to have been formed by bulk sediment processing at the
anterior of the burrow during continuous burrowing rather than successive
probing as is proposed for Phycosiphon s.s.
|