|
|
|
Results
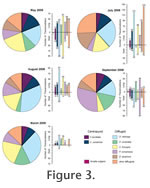
The dominant centropyxid taxon in May and July was Centropyxis constricta (Ehrenberg, 1843), whereas Centropyxis aculeata (Ehrenberg, 1832) increased in abundance during August and September (Figure 3). Arcella vulgaris Ehrenberg, 1830 was present in low numbers in May and increased in numbers in September. Cucurbitella tricuspis (Carter) Medioli et al., 1987 remained an important component of the thecamoebian population from May to August. In August the highest proportion of the C. tricuspis population was alive, while in September its numbers decreased significantly. The relative abundance of difflugiid (vs. centropyxid) thecamoebians comprising the monthly populations remained relatively consistent over the course of the study (ranging from 74.6 to 87.8%) (Table 1 and Figure 3). However, the thecamoebian taxa composing the difflugiid population varied greatly, with P-values > 0.05. Difflugia oblonga Ehrenberg, 1832 remained relatively ubiquitous throughout the study (Figure 3), its living population peaked during the warmest months of July and August, then its total numbers decreased significantly in September. Difflugia urceolata (Carter, 1864) was present (stained and empty tests) during both May and September but was virtually absent in July and August. Substantial increases of both D. urceolata and Pontigulasia compressa (Carter, 1864) occur in the month of September (Figure 3). The thecamoebians composing the group "Others" in Figure 3 are Lagenodifflugia vas (Leidy, 1874), Difflugia protaeifomis Lamarck, 1816, Difflugia bidens Penard, 1902, Diffligia corona Wallich, 1864, Difflugia bacillaliarum Perty, 1849 and Difflugia globula (Ehrenberg, 1848). The proportion of the species grouped as "Others" remains relatively consistent throughout the study, except for the month of July. An increase in the number of "Others" in July is due to an increase in the number of living D.globula. The dominant thecamoebian in July was D. amphora. The thecamoebian taxa observed in the month of March are common species observed in the fossil record with very few stained tests (Boudreau et al., 2005; McCarthy et al., 1995) (Figure 3). The majority of the population observed in March was composed of encysted thecamoebians |
|