|
|
|
MATERIALS AND METHODSFossil Material
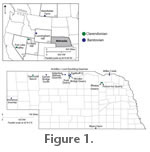
The largest sample of Hesperolagomys studied, representing a previously undescribed species, derives from three of the five Barstovian-age Valentine Railway Quarries (Valentine Quarry B, West Valentine Quarry, and Stewart Quarry) in the Crookston Bridge Member of the Valentine Formation in north-central Nebraska. Important specimens of Hesperolagomys (sp. nov.) from the previously unpublished Garner Bridge South faunal assemblage are included in this study. Specimens referred to Hesperolagomys from the Barstovian Norden Bridge Quarry Fauna and Nebraska faunas of equivalent age from the Cornell Dam Member of the Valentine Formation (Achilles, Lost Duckling, and Egelhoff Quarries) were studied (Voorhies 1990a, 1990b). The hypodigm and previously undescribed topotypes of H. galbreathi from Fish Lake Valley were included in this study. Well-preserved specimens referable to H. galbreathi from the Tiensvold Ranch faunal assemblage, in Ash Hollow Formation sediments in Sheridan County of northwestern Nebraska, were also studied. Other Hesperolagomys specimens of Clarendonian age from Nebraska included here are from Poison Ivy Quarry, northeastern Nebraska, and from Pratt and Bluejay Quarries in the Merritt Dam member of the Ash Hollow Formation of north-central Nebraska (Korth 1998). Clarendonian Hesperolagomys described from Scrappy Hill, Utah (Tedrow and Robison 1999), are also included in this study. The hypodigm and previously undescribed topotypes of Hesperolagomys fluviatilis from the Barstovian Kleinfelder Farm locality, Wood Mountain Formation (Storer 1970, 1975) were studied. Barnosky (1986) described Oreolagus colteri (referred below to Hesperolagomys sp.) from the Barstovian Cunningham Hill Fauna, Colter Formation, western Wyoming. Comparative MaterialSpecimens included in this study were compared with specimens of other lagomorphs including the extinct ochotonids Russellagus vonhofi, Oreolagus nebrascensis, Gripholagomys lavocati, Amphilagus fontannesi, Amphilagus ulmensis, and the extinct leporids Palaeolagus, Megalagus, Archaeolagus, and Hypolagus in the collections of the University of Nebraska State Museum (Division of Vertebrate Paleontology), Royal Ontario Museum, University of Michigan Museum of Paleontology, Museum of Paleontology at the South Dakota School of Mines and Technology, University of California Museum of Paleontology, and the Carnegie Museum of Natural History. Comparison was made with the published description of Cuyamalagus dawsoni (Hutchison and Lindsay 1974). Specimens of extant Ochotona princeps (adult skulls: ZM 2114, 2117, 2118, 14366; adult skeleton: ZM 2114; juvenile skull: ZM 2134), Sylvilagus audubonii (adult skulls: ZM 3281 and 3284) and Sylvilagus floridanus (adult skulls: ZM 12086, 12087, 17159; juvenile skull: ZM 26135) in the collections of the Division of Zoology, University of Nebraska State Museum, were also used for comparison. Methods
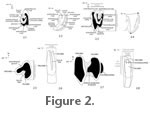
Crown height of p4 (the lower cheek tooth with the highest crown) was measured on the trigonid from crown apex to the base of the lingual hypostria (see Figure 2.8). Hypsodonty indices for p4 were calculated as crown height divided by anteroposterior length using unworn or nearly unworn teeth (possessing a posterolophid completely isolated from the main body of the talonid). Because upper cheek teeth are strongly buccolingually curved, straight-line measurements severely underestimate crown height. I devised a new method to measure the height of the curved crown of P4s, the upper cheek teeth with the highest crowns (as shown in Figure 2.4). Scaled camera lucida line drawings or digital photographs of the lingual margin (tallest part of the crown), in anterior or posterior view (which give equivalent measures of crown height for P4s), were input into Object-Image (Vischer 2003), which was used to calculate the length of the curved line representing the crown height. I also used this technique to measure tip-to-base length of incisors. Upper cheek teeth demonstrate considerable delay in completion of the crown and root until after occlusal wear has occurred ('incipient hypselodonty', e.g., Webb and Hulbert 1986), such that the 'complete' crown height and degree of hypsodonty cannot be measured directly from any single tooth. To obtain an estimate of the 'complete' crown height of P4 I overlaid line drawings of the lingual profiles of teeth with unworn or nearly unworn crowns and more heavily worn teeth with completed crowns, and measured the length of the curved line representing the 'complete' crown using the methodology described above for measuring crown height of P4 of individual specimens. I calculated an estimate of the hypsodonty index of P4 as the 'complete' crown height divided by the mean value of anteroposterior length. To determine the significance of the relationship between crown height (as a proxy for stage of wear) and occlusal dimensions in P4 of Hesperolagomys, I tested the linear correlation of crown height with width and length of P4s from the largest (Valentine Railway Quarries) sample. For this analysis, I used only those crown height values measured directly from specimens, omitting the estimated maximum crown height values obtained by the method described above. I calculated minimum, maximum, and mean values, standard deviation and coefficient of variation for each tooth dimension for specimens of H. fluviatilis from the Kleinfelder Farm locality sample and a combined Valentine Railway Quarries and Garner Bridge South sample of Hesperolagomys. I also tested tooth dimensions of the Valentine Railway Quarries and Garner Bridge South pooled sample to determine if distributions approximated normality using Shapiro-Wilk W tests. One-way ANOVAs on length of P3-M2, length and width of p3, and length and trigonid width of p4-m2 were performed to evaluate the significance of size differences between species. An additional ANOVA was performed using a pooled sample of m1s and m2, as these teeth are sometimes difficult to differentiate in practice. Hesperolagomys sp. nov. and H. fluviatilis samples from the combined Valentine Railway Quarries + Garner Bridge South and Kleinfelder Farm, respectively, as described above were included, and a pooled sample of H. galbreathi was composed of topotypes from Fish Lake Valley and specimens from the Tiensvold Ranch locality. Preliminary analysis of specimens from the two Clarendonian localities revealed that they could not be distinguished morphologically or statistically. Pairwise comparisons were conducted using Student's t tests (Zar 1999; Sall et al. 2005). I also calculated estimates of power of the ANOVAs to investigate if small sample size might reduce the meaningfulness of the analyses; I varied sample size from that of the actual sample to 100 in increments of 1. AbbreviationsAnatomical Abbreviations. ant, anterior; a-p, anteroposterior; CH, crown height; L, length; lin, lingual; tr, transverse; tri, trigonid; tal, talonid; W, width. Institutional Abbreviations. IMNH, Idaho Museum of Natural History; MCZ, Harvard Museum of Comparative Zoology; ROM, Royal Ontario Museum; UCMP, University of California Museum of Paleontology; UNSM, University of Nebraska State Museum; UWBM, University of Washington-Burke Museum. |
|