1La conservation de la biodiversité est devenue aujourd’hui un impératif pour toute société s’inscrivant dans l’optique du développement durable. Cette conservation doit reposer sur une bonne connaissance de la diversité biologique pour être efficiente et mener des actions adéquates (Ouoba, 2006). La convention sur la diversité biologique élaborée à Rio de Janeiro en 1992 a souligné la nécessité de prendre des mesures adéquates pour la protection et la conservation des écosystèmes. Plusieurs États africains ont ainsi entrepris de créer de nouvelles aires protégées sur leur territoire (UICN, 1994). Au Togo, la nécessité de conserver les ressources naturelles remonte à l’époque précoloniale avec les aires sacrées, présentées comme des stratégies locales de conservation de la biodiversité (Kokou & Sokpon 2006). À l’époque coloniale, de nombreuses aires protégées ont été créées à travers le pays, dont la réserve de l’Oti-Mandouri. Depuis son accession à l’indépendance, le Togo a pris conscience de la nécessité de protéger ses richesses biologiques en adoptant sur le plan national des stratégies, des plans d’action et des politiques sectorielles.
2En Afrique de l’Ouest, la faible implication des populations riveraines dans la gestion des aires protégées conduit fréquemment au non-respect des règlements en vigueur (Wafo Tadopda, 2011), qui se traduit par l’envahissement de ces espaces. Au Togo, c’est au début des années 90, suite aux troubles socio-politiques qu’a connus le pays, que ces envahissements ont pris de l’ampleur (SOTED, 2001).
3Aujourd’hui, la réserve de l’Oti-Mandouri est devenue la cible privilégiée des exploitants de bois, des agriculteurs, des pêcheurs et des éleveurs. Sous la pression de leur croissance démographique conjuguée à une paupérisation grandissante, les populations riveraines violent ce domaine protégé à la recherche de nouvelles terres pour l’agriculture ou des coupes pour le bois de chauffe. Ces activités contribuent à la dégradation alarmante des écosystèmes de cette aire protégée, notamment des changements majeurs dans la structure, la composition floristique et la productivité de ces formations végétales. Cette situation enlève à cette aire protégée son rôle de protection et de conservation in situ de la diversité biologique ; d’où la nécessité de mener des actions pour une protection véritable à travers un plan d’aménagement et de gestion durable. Ces actions doivent reposer sur des données de base portant sur la diversité biologique et sa distribution dans la réserve l’Oti-Mandouri actualisées régulièrement. Or, celles-ci font actuellement défaut.
4L’intégration de la recherche aux stratégies de gestion des aires protégées doit leur permettre d’assurer un soutien scientifique permettant de les asseoir dans la stratégie globale de gestion efficiente de l’environnement et de la diversité biologique. C’est dans cette optique que la présente étude est menée afin de collecter et analyser les données de base pour contribuer à la gestion durable de la réserve de l’Oti-Mandouri. Il s’est agi de déterminer la diversité floristique et la structure des différentes formations végétales; d’identifier les formes d’exploitation des ressources végétales de cette réserve et de cartographier les traces d’activités humaines à l’intérieur de la réserve.
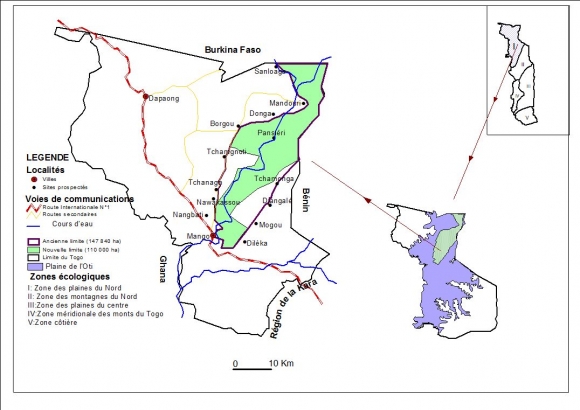
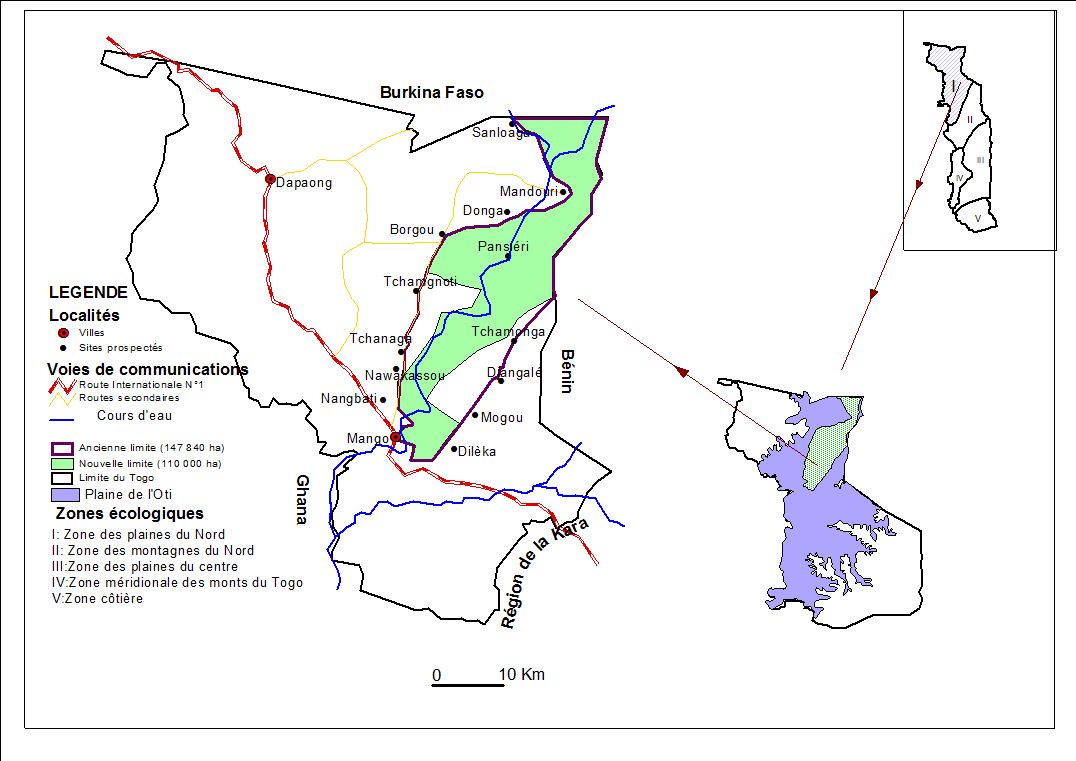
5L’étude est menée dans la réserve de l’Oti-Mandouri située dans la région des Savanes entre 0°30’ et 0°47’ de Longitude Est et entre 10°18’ et 11° de latitude Nord. La zone d’étude est localisée dans la zone écologique I (Ern, 1979) qui correspond aux plaines du nord, couvertes principales de savanes sèches. Sur le plan phytogéographique, elle est située dans le centre Régional d’endémisme soudanien (White, 1986).
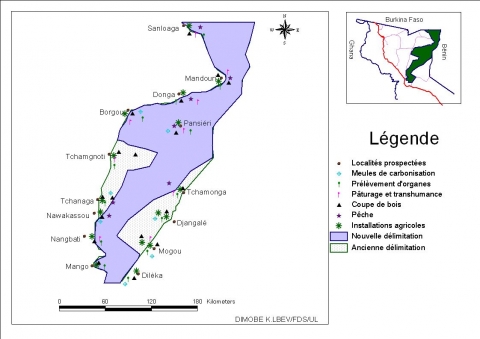
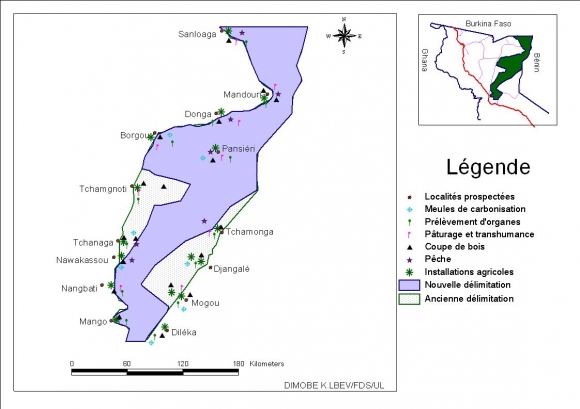
6Cette réserve fait frontière commune avec le Burkina Faso sur 22 km au Nord et avec le Bénin sur 46 km à l’Est. L’ancienne route Mango-Borgou-Mandouri constitue sa limite Ouest. Elle fut classée en 1981 pour servir de couloir de migration des grands mammifères qui se déplacent annuellement entre le parc national de l’Oti- Kéran au Togo et les parcs nationaux d’Arly (Burkina Faso) et de la Pendjari (Bénin). Ces limites ont été repoussées à plusieurs reprises portant sa superficie à 147 840 ha, dont les trois quarts appartiennent à la préfecture de l’Oti et le quart à celle de Kpendjal. Aujourd’hui, 37 840 ha situés en périphérie ont été déclassés (dû aux envahissements) et la superficie totale de la réserve n’est plus que de 110000 ha (Figure 1), (UICN/PACO, 2008). Le relief est marqué par les plateaux de Dapaong-Bombouaka et par la plaine de l’Oti dont l’extrême nord-est fait l’objet de la présente étude.
Figure 1. Localisation de la zone d’étude
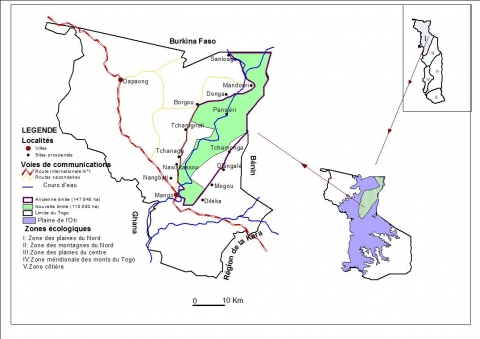
7La plaine de l’Oti a des altitudes basses (120 à 200 m). Elle est caractérisée par la monotonie d’ensemble de son paysage. Mais dans le détail, elle est formée d’une succession de bas plateaux très faiblement inclinés vers les talwegs qu’ils dominent d’une vingtaine à une trentaine de mètres. Les sols rencontrés dans la zone sont essentiellement ferrugineux tropicaux de type latéritique encroûté, des sols peu évolués et des sols hydromorphes le long de la rivière Oti. Le régime pluviométrique est caractéristique des climats tropicaux avec l’alternance de deux saisons : une saison de pluie de cinq mois (mai à octobre) et une saison sèche de sept mois durant le reste de l’année. Les températures varient entre 17 et 39 °C en saison sèche et entre 22 et 34 °C en saison des pluies. Mango, une des grandes villes au cœur de la plaine de l’Oti enregistre le maximum absolu de température (41 °C) pendant le mois d’avril. La pluviométrie varie entre 800 à 1 100 mm.
8La réserve de l’Oti-Mandouri est marquée par une forte emprise des activités anthropiques sur les forêts galeries et les savanes qui sont les formations végétales dominantes. Les principales activités sont l’agriculture, l’élevage, la collecte de bois de chauffe et la pratique des feux de brousse en saison sèche. Le peuplement humain est constitué essentiellement de quatre groupes ethniques. Il s’agit des Moba et Gourman représentant 63 %, les Tchokossi 10 % et les Ngan-Ngan 7 %. Les Tchokossi sont concentrés dans la préfecture de l’Oti, les Gourman et Moba dans la préfecture de Kpendjal (Gayibor, 1997). Les mouvements migratoires vers d’autres régions pour l’agriculture sont fréquents ; la faible fertilité des sols couplée probablement à la densité de population en serait la cause. L’agriculture et l’élevage constituent les principales activités pour les populations qui en tirent leurs revenus. Les principales cultures sont le sorgho, le mil, le fonio, l’arachide, le niébé, le maïs, l’igname, le manioc, etc.
9Les relevés floristiques ont été effectués le long de transects transversaux de l’extérieur vers l’intérieur de la réserve sur des aires floristiquement homogènes dans lesquelles 126 placeaux d’inventaire ont été installés. Dans les savanes et forêts claires, les placeaux avaient une aire de 900 m2 (30 m x 30 m). Le long des cours d’eau, les placeaux avaient une forme plus linéaire (50 m x 10 m) en vue d’épouser le réseau hydrographique (Natta et al, 2003).
10Dans chaque placeau d’inventaire, trois placettes de régénération de 5 m x 5 m ont été installées. L’inventaire floristique a consisté à relever toutes les espèces ligneuses présentes dans les placeaux, en leur affectant un coefficient d’abondance-dominance suivant l’échelle de Braun-Blanquet (1932). Les espèces non identifiées sur le terrain ont été récoltées, déterminées et confirmées au Laboratoire de Botanique et Écologie Végétale de l’Université de Lomé à partir des flores de Hutchinson et Dalziel (1954-1972), Brunel et al. (1984), Keay & Hepper (1954-1972), Scholz & Scholz (1983), Akoègninou et al. (2006). Une fiche standard de descripteurs écologiques était remplie en même temps que les relevés floristiques étaient réalisés. Dans chaque placeau, toutes les espèces d’arbres ont été relevées et pour tous les individus, les paramètres dendrométriques (hauteur, diamètre mesuré à 1,30 m au-dessus du sol pour les individus à dbh ≥ 10 cm) ont été mesurés. Les coordonnées géographiques des placeaux d’inventaire floristique et des traces d’activités humaines ont été enregistrées à l’aide du GPS (Global Positioning System).
11L’analyse de la relation entre les populations riveraines et la réserve d’une part, et entre les populations et les agents forestiers d’autre part a été faite par une enquête auprès des acteurs en présence. Cette dernière a été effectuée sous forme d’entretien semi-structuré (Mary & Besse, 1996). Ces entretiens reposaient sur un questionnaire testé au préalable (Martin, 1995). Deux types de questionnaire ont été élaborés à cet effet et administrés respectivement aux populations locales et aux gestionnaires. Les questionnaires comportaient des variables d’identification des enquêtés (nom, prénoms, âge, sexe, niveau d’instruction, village d’origine) et des questions relatives aux activités pratiquées à l’intérieur et à la périphérie de la forêt, à la perception que les populations ont de la réserve, de leur participation à la gestion de cette aire protégée ainsi que des difficultés rencontrées par les agents forestiers dans l’accomplissement de leur mission.
12Afin de s’assurer de la bonne formulation des questionnaires, ces derniers ont été administrés à titre d’essais à quelques villageois choisis au hasard dans le milieu d’étude. Ceci nous a permis d’enlever des questions inutiles et d’en rajouter d’autres.
13Au cours de cette étude dix-huit villages riverains ont été prospectés et 101 personnes ont été enquêtées. Les enquêtes ont été complétées par des observations directes aux champs et dans les formations naturelles (Twamasi, 2001; Tom Kumekpor, 2002) avec l’aide des agents forestiers ou des guides locaux. Cette prospection des formations naturelles a permis d’identifier sur le terrain les principales activités illicites menées dans la réserve et les espèces exploitées. Les observations sont consignées dans un carnet de notes. Des photos ont été prises sur le site de même que des coordonnées géographiques des relevées à l’aide d’un récepteur GPS à des fins d’analyse spatiale. Les membres des mouvements associatifs de la zone d’étude ont été interrogés. Il s’agit du président de l’UAVGAP (Union des Associations Villageoise de Gestion participative des Aires Protégées) à Mango ; les membres du comité des AVGAP (Association Villageoise de Gestion participative des Aires Protégées), les sages et les populations riveraines de la réserve et toute autre personne ressource disponible et susceptible de nous fournir des informations.
14La richesse spécifique, les fréquences et abondances spécifiques ont été calculées et des diagrammes rang-fréquences (Magurran, 1988) ont été réalisés. De plus, une matrice «relevés x espèces» a été élaborée sur la base de la présence/absence des espèces, avec en ligne les espèces et en colonnes les relevés. À chaque espèce relevée, la famille, la forme biologique (Raunkaier, 1934) et l’affinité phytogéographique (White, 1986) correspondantes ont été affectées.
15La matrice élaborée a été soumise à des techniques d’analyse multivariée dans le but de mettre en évidence les gradients écologiques majeurs ainsi que les principales formations végétales qui s’en dégagent. Parmi la multitude de techniques qui existent, la méthode de TWINSPAN® (Two Way INdicator SPecies ANalysis) (Hill, 1979) a été choisie pour la description physionomique des classes de végétation. La méthode de DECORANA® (DEtrended CORrespondance ANAlysis) a été utilisée pour l’ordination des relevés. Ces deux analyses ont été réalisées à l’aide du logiciel CAP® (Community Analysis Package).
16La détermination de la diversité alpha a été faite par le calcul de la richesse spécifique (Rs), de l’indice de diversité de Shannon (H’), de l’équitabilité de Pielou (E) (Magurran, 1988 ; Kent et Coker, 1992).
17Les types biologiques permettent une appréciation très fructueuse des phytoclimats et sont très précieux pour la recherche de leur évolution écologique (Trochain, 1966). Les types biologiques considérés pour l’analyse des spectres biologiques sont ceux définis par Raunkiaer (1934), repris et aménagés par diverses études de la végétation des régions tropicales (Breman, 1978 ; Lebrun, 1981 et Aké Assi, 1984). Il s’agit de phanérophytes (Ph), chaméphytes (Ch), Hémicryptophytes (He), géophytes (Ge), thérophytes (Th). Les formes lianescentes (Lmph, Lnph, LmPh, LnPh) ont été aussi distinguées. Les phanérophytes se composent de :
-
mégaphanérophytes (MPh) : arbres de plus de 30 m de haut ;
-
mésophanérophytes (mPh) : arbres de 10 à 30 m de haut ;
-
microphanérophytes (mph) : arbres de 2 à 10 m de haut ;
-
nanophanérophytes (nph) : arbres de à 0,4 à 2 m de haut ;
18D’après Aubréville (1963), les spectres biologiques, à savoir le spectre brut et le spectre réel sont « des notions caractéristiques des formations végétales en corrélation directe avec le milieu ». Dans le présent travail, le spectre brut est considéré, le plus utilisé. Il est déterminé par le pourcentage du nombre d’espèces correspondant à chaque type biologique, par rapport au nombre total d’espèces. Il permet de mettre en évidence la signification floristique de chaque type biologique et de chaque strate dans les différentes formations.
19Les subdivisions chorologiques utilisées dans l’analyse des spectres phytogéographiques sont celles définies par (White, 1986). Il s’agit des :
-
-
espèces à large distribution comme les espèces afro-américaines (AA), pantropicales (Pan), paléotropicales (Pal) ;
-
espèces à distribution continentale regroupant les espèces soudano-guinéennes (SG), afro-tropicales (AT), afro-malgaches (AM), pluri-régionales africaines (PRA), soudano-zambéziennes (SZ) et guinéo-congolaises (GC) ;
-
espèces de l’élément-base soudanien (S) distribuées dans le Centre Régional d’Endémisme (CRE) soudanien.
20Le traitement des données d’enquête collectées sur le terrain en vue de la rédaction de cet article s'est effectué manuellement à l'aide d'une grille de dépouillement. Ce dépouillement a permis d'apprécier la fréquentation de la réserve de l’Oti-Mandouri par les populations et les impacts de leurs activités sur la préservation des ressources de l'aire protégée. L’analyse spatiale des pressions anthropiques sur la réserve de l’Oti-Mandouri a été abordée par la mise en place d’un SIG simplifié sur les traces d’activités notées dans ladite réserve. Les coordonnées géographiques des placeaux d’inventaire et des traces d’activités enregistrées au GPS lors de la prospection sur le terrain ont été projetées sur un fond de carte géoréférencée de la réserve de l’Oti-Mandouri à l’aide du logiciel Arcview®. Ceci a permis de réaliser la carte de distribution des traces d’activités humaines à l’intérieur ou à la périphérie de la réserve.
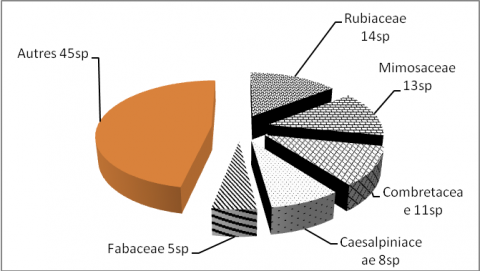
21Les investigations botaniques menées dans la réserve de l’Oti-Mandouri ont permis de compter et mesurer dans 126 placeaux 925 individus appartenant à 116 espèces ligneuses. Les espèces inventoriées se répartissent en 84 genres et 33 familles (Annexe). Les trois familles les plus représentées (38,77 % des espèces) ont plus de dix espèces chacune. Il s’agit des Rubiaceae (14,28 %), des Mimosaceae (13,26 %) et des Combretaceae (11,22 %) (Figure 2).
Figure 2. Spectre spécifique des familles (sp : espèces)
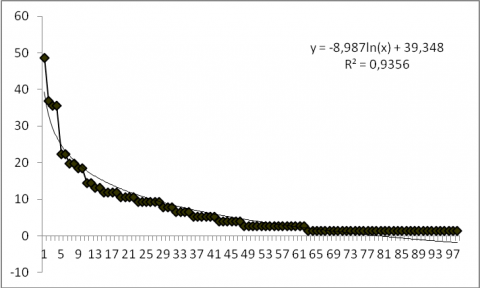
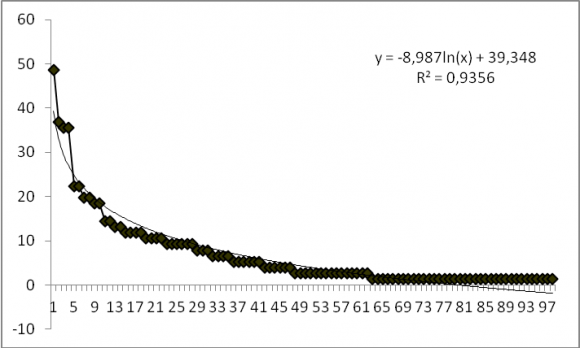
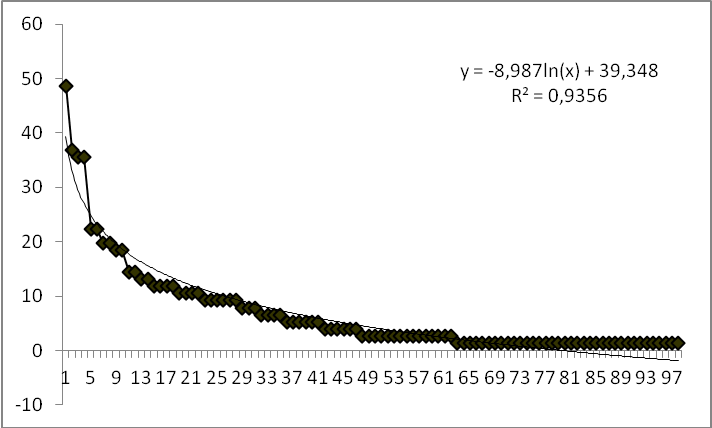
22La courbe de distribution des fréquences spécifiques présentant une allure décroissante bien ajustée à une fonction logarithmique illustre bien ce résultat (Figure 3). Combretum glutinosum (48,68 %) présente une fréquence relative de plus de 40 %, d’où la dominance de cette espèce dans la réserve de l’Oti-Mandouri. Quatre (4) espèces (4,08 %) ont une fréquence relative > 30 %. Il s’agit de Combretum glutinosum (48,68 %), Mitragyna inermis (36,84 %), Acacia polyacantha (35,52 %) et Piliostigma thonningii (35,52 %). Vingt-sept (27) espèces ont une fréquence relative comprise entre 7 et 28 % (Pterocarpus erinaceus, Terminalia avicinoides, Paulinia pinnata, Prosopis africana, Nauclea latifolia, Terminalia macroptera, Gardenia ternifolia, Pterocarpus santalinoides, Acacia gourmaensis etc). Enfin quatre-vingt-cinq (85) espèces ont des fréquences relatives inférieures ou égales à 6 %.
Figure 3. Distribution des fréquences spécifiques en fonction du rang
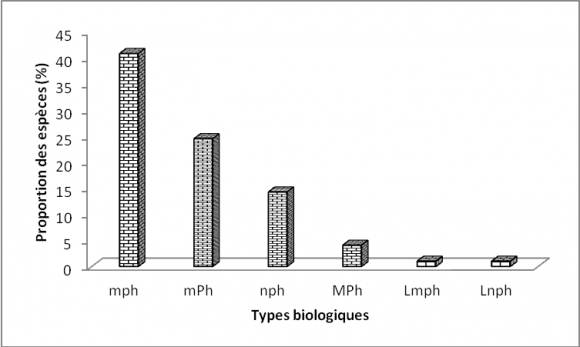
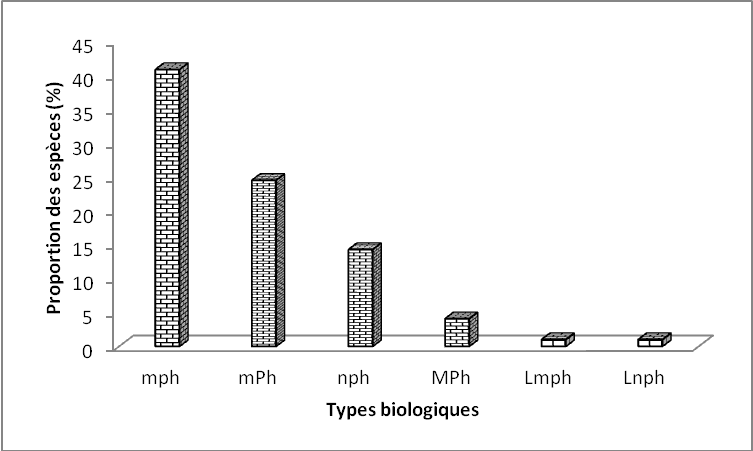
23Les microphanérophytes (40,81 %) sont les plus représentées, suivies des mésophanérophytes (24,48 %), des nanophanérophytes (14,28 %) et des mégaphanérophytes (1,63 %).
24Les lianes regroupent les microphanérophytes lianescentes (Lmph=1,02 %) et les nanophanérophytes lianescents (Lnph=1,02 %) (Figure 4).
Figure 4. Spectre des types biologiques
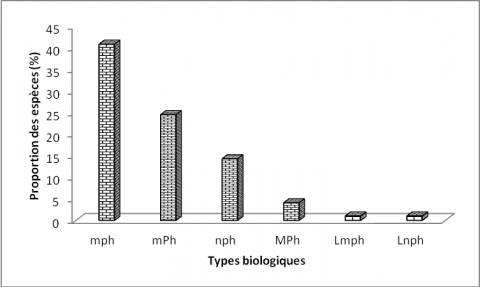
Légende :¨mp : microphanérophytes, mP : mésophanérophytes, np : nanophanérophytes, Ch : chaméphytes, He : hémicryptophytes, Ge : géophytes, Th : thérophytes, Lmp : mésophanérophytes lianescents, Lnp :nanophanérophytes lianescents
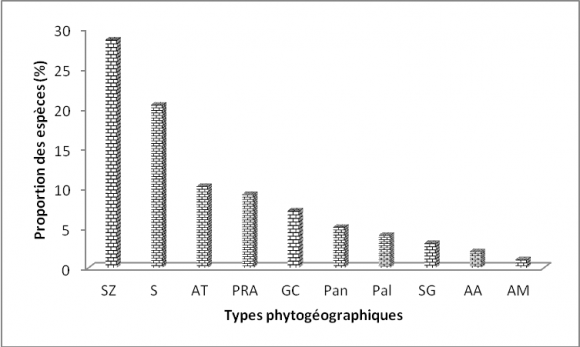
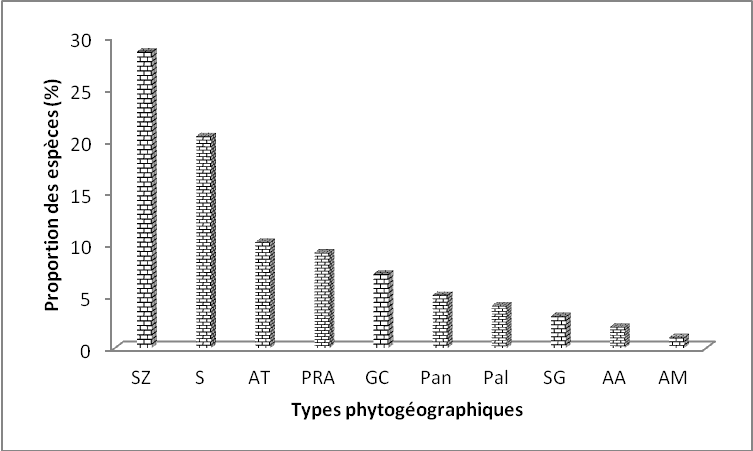
25La florule de la réserve de l’Oti-Mandouri est dominée par les espèces soudano-zambiennes (28,57 %) suivies des espèces de l’élément base-soudanien (20,40 %), les espèces afro-tropicales (10,20 %) et les espèces pluri-régionales africaines (9,18 %). Les espèces guinéo-congolaises (7,14 %), pantropicales (5,10 %), paléotropicales (4,05 %), soudano-guinéennes (3,06 %) afro-asiatiques (2,04 %) et afro-malgaches (1,02 %) sont peu représentées (Figure 5).
Figure 5. Spectre global des types phytogéographiques
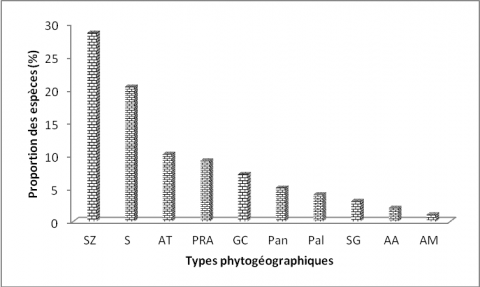
Légende :¨SZ : soudano-zambézien, GC : guinéo-congolaises, S : élément-base soudanien, AT : afro-tropicales, Pan : pantropicales, PRA : plurirégionales africaines, SG : soudano-guinéennes, Pal : paléo-tropicales, AM : afro-malgaches, AA : afro-asiatiques)
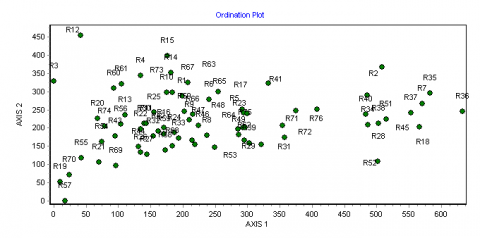
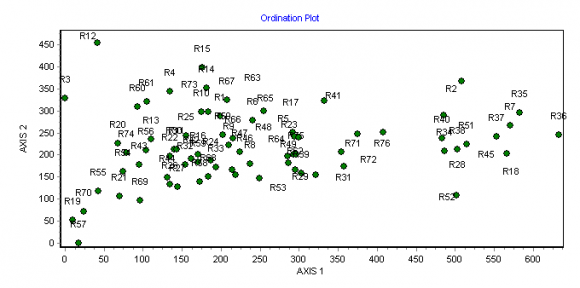
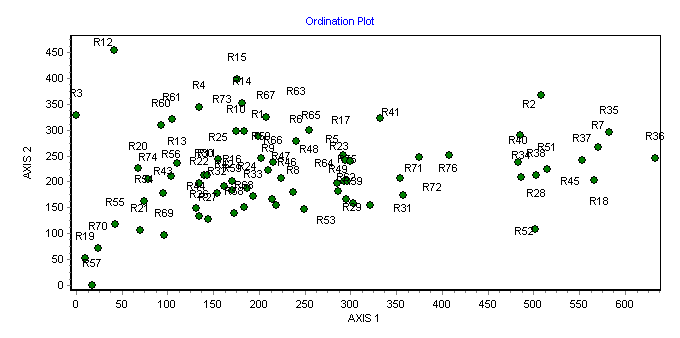
26Les analyses multivariées ont permis de discriminer les principales formations végétales et de mettre en évidence les gradients écologiques de leur distribution. En effet, la disposition des placeaux d’inventaire dans le plan factoriel des axes 1 et 2 de l’ordination des formations échantillonnées révèle l’influence d’un gradient écologique (Figure 6). L’axe 1 présente une succession de la droite vers la gauche, des forêts galeries (FG) le long des cours d’eau, les savanes boisées (SB) dans les zones inondables, les savanes arborées (SA) et savanes arbustives (Sa). Cet axe traduit un gradient édaphique et discrimine les formations des sols peu profonds et gravillonnaires (vers la gauche) de ceux des sols profonds et argileux (vers la droite). Le même axe indique que l’humidité relative du sol augmente à mesure que l’on passe de la gauche vers la droite. Ce gradient d’humidité induit aussi un gradient de fermeture ou de densité de la végétation, étant donné que sont séparées de la gauche vers la droite du graphique les formations plus ou moins fermées.
Figure 6. Diagramme de l’ordination des placeaux dans le plan factoriel des axes 1 et 2 de la DCA
27Au total, neuf types d’activités anthropiques menées dans la réserve de l’Oti-Mandouri ont été recensées, qui sont autant de formes de pressions sur les ressources végétales de cette aire protégée. Six d’entre eux relèvent des activités extractivistes qui s’exercent soit sur la faune, il s’agit de la chasse et de la pêche, soit sur la flore : prélèvement de plantes médicinales, coupe et ramassage de bois, carbonisation, prélèvement de paille dans la savane pour la confection des toits des cases. Trois d’entre eux relèvent de l’occupation de l’espace protégé et de sa transformation à des fins anthropiques, il s’agit a) des feux de brousse effectués dans le cadre de l’écobuage pour favoriser le pacage du bétail ou l’activité agricole, b) de la mise en culture, c) de la transhumance des troupeaux.
28La pêche s’effectue dans la réserve bordure de la rivière de l’Oti pendant la saison sèche. (Figures 7 et 8). Il n'y a pas une mare, une rivière qui ne soient zébrées de petits barrages de décrue pour la capture du poisson. La rivière est, quant à elle, sillonnée par les pirogues. La pêche (bien qu’interdite dans la réserve) sur la rivière Oti est pratiquée d'une manière anarchique avec l'utilisation des engins de pêche non règlementaires tels que les sennes et les éperviers avec des mailles inférieures à 20 cm de diamètre (MERF, 2002). Les enquêtes effectuées auprès des pêcheurs ont montré que ce système d’exploitation des ressources et les diverses techniques ont conduit à un appauvrissement soutenu des cours d’eau. À cette pression halieutique se surimposent d’autres formes de perturbation des cours d’eau. Il s’agit en premier lieu de leur comblement partiel par le sable mis en suspension par les défrichements. En second lieu, il s’agit des perturbations chimiques ou biologiques occasionnées par l'utilisation de pesticides et engrais minéraux sur les cultures de coton ou les zones maraîchères, particulièrement développées sur les rives. Au final, toutes ces perturbations se conjuguent pour entraîner une baisse de l’abondance de poisson. Ainsi 71,85 % des pêcheurs que nous avons enquêté nous ont déclaré qu'ils n'arrivent plus à attraper de gros poissons et en quantité suffisante.
Figure 7. La pêche au filet sur Oti

Photo : K. Dimobe
Figure 8. Produit de pêche à Tchamgnoti

Photo : K. Dimobe
29Dans toute aire protégée, la chasse est synonyme de braconnage. Dans la réserve de faune de l’Oti-Mandouri, tous les points d'eau isolés sont accompagnés d'une petite hutte ou d'un abri de branchages utilisés par les chasseurs. Les entretiens tenus avec des chefs de ménages et des chasseurs nous ont permis de cerner l'origine et les causes du braconnage, dont le développement obéit à trois causes principales :
-
-
un accès facile aux armes à feu ;
-
une grande demande pour la viande de brousse et la commercialisation des produits de la chasse ;
-
un bas revenu et peu d'opportunités pour les populations périphériques.
30Face à ces facteurs favorables au braconnage, les réponses des gestionnaires de la réserve sont des plus limitées : les moyens humains sont trop réduits (sept agents forestiers pour une zone de 110 000 ha), les moyens matériels de surveillance sont également insuffisants (trois motos). Il n’est donc pas étonnant qu’aient lieu des intrusions irrégulières des populations à l'intérieur du parc, occasionnant un braconnage intensif.
31La transhumance à l’intérieur de la réserve des troupeaux de bovins en provenance des pays sahéliens est un phénomène généralisé (Figure 9 et 10). L'effectif du cheptel ne cesse d'augmenter chaque année et aucun endroit de la réserve n’est épargné. Cette situation pourrait s'expliquer par:
-
-
la bonne maîtrise des différentes pathologies animales
-
La disponibilité du fourrage avec des points d'eau permanents dans la réserve qui constituent autant de points d'attraction pour les animaux domestiques.
Figure 9. Transhumance à Nangbati

Photo : K. Dimobe
Figure 10. Transhumance à Borgou

Photo : K. Dimobe
32Le cheptel présent doit représenter quelques centaines de têtes. Tous les pâturages, les abords de mares, les points d'eau, les rives de la rivière Oti montrent des traces nombreuses de passage de ces animaux. Au voisinage des nombreux campements d'éleveurs, les espèces végétales émondées par la nourriture des troupeaux témoignent de l'impact négatif de la pression pastorale sur les ressources naturelles de la réserve. Les espèces végétales consommées par le bétail sont : Afzelia africana (menacée d’extinction dans la zone à cause de sa fréquence d’utilisation), Prosopis africana, Pterocarpus erinaceus, Khaya senegalensis, Stereospermum kunthianum. Au final, la présence de bétail dans la réserve constitue une menace pour les écosystèmes et les espèces en raison de la perturbation de la faune et de la flore, de la compétition de la faune sauvage et du bétail pour les ressources alimentaires, des risques de transmission d'épizooties à la faune sauvage et des risques d'empoisonnement des grands carnivores par les éleveurs.
33La carbonisation, ou production de charbon de bois (Figure 11 et 12) n’est assurée que par un nombre limité d'exploitants pour qui cette activité est généralement secondaire. Les espèces utilisées dans la fabrication du charbon de bois sont : Prosopis africana (cité par 82,05 % des enquêtés), Anogeisus leiocarpus (75,22 %), Pterocarpus erinaceus (60,17 %), Terminalia spp. (26,79 %), Khaya senegalensis (11,32 %).
Figure 11. Meule de carbonisation à Dilèka

Photo : K. Dimobe
Figure 12. Sacs de charbon prêts pour la vente à Pansiéri

Photo : K. Dimobe
34Les meules de carbonisation sont situées à l’intérieur de la réserve (Figure 7).
35Le prélèvement des plantes médicinales : Les résultats d’enquête et les observations faites sur le terrain (Figurs 13 et 14) ont permis de constater que les populations riveraines utilisent les produits forestiers non ligneux (racines, écorce et feuilles) dans la pharmacopée traditionnelle (pour se soigner).Aussi, la demande en remèdes traditionnels dans la zone d’étude a enregistré une hausse du fait de la croissance démographique et du coût élevé de la médecine occidentale. Ces informations ont été données par des personnes ressources (les guérisseurs traditionnels) lors des enquêtes. Les espèces fréquemment prélevées à des fins médicinales sont : Sclerocarya birrea (cité par 56,67 % d’enquêtés), Nauclea latifolia (44,98 %), Azadirachta indica (40,10 %), Cassia sieberiana (38,12 %), Xeroderris sthulmanii (23,67 %), Pteleopsis suberosa (22,77 %), Khaya senegalensis (21,11 %).
Figure 13. Prélèvement d’écorce de Sclerocarya birrea

Photo : K. Dimobe
Figure 14. Prélèvement d’écorce de Sterculia setigera

Photo : K. Dimobe
36Selon la Figure 15, les prélèvements d’organes se font un peu partout dans la réserve.
Figure 15. Carte de distribution spatiale des activités humaines dans la réserve de l’Oti – Mandouri
La coupe de bois et le ramassage de bois: La coupe du bois est omniprésente. Les souches fraîchement coupées sont bien visibles (Photo 9) pratiquement partout dans l’aire protégée, et ce même loin des limites externes. De nombreux tas de bois en attente d'être emportés sont visibles (Photo 10). Le bois de feu est la principale source d'énergie dans la zone d’étude. Il est utilisé dans les foyers pour la cuisson des aliments. Il est essentiellement collecté par les femmes. À cause des revenus qu'il génère, certains hommes s'adonnent à cette activité (et 17). Mais avec l'augmentation des besoins (domestiques et commerciaux), les ressources en bois au niveau des jachères deviennent insuffisantes. La durée des jachères a considérablement diminué devant l'extension des zones mises en cultures en relation avec la pression démographique croissante. D'après les enquêtes, les espèces les plus appréciés comme bois de feu sont Anogeissus leiocarpus (78,86 %), Prosopis africana (71,25 %), Pterocarpus erinaceus (57,81 %), Terminalia macroptera (39,15 %), Gardenia ternifolia (13,51 %), etc. parce qu'elles sèchent très vite, conservent la flamme, prennent feu rapidement et ne font pas trop de fumée.
Figure 16. Souche de Prosopis africana coupée

Figure 17. Tas de bois en attente pour la vente

Figure 18. Ramassage de bois pour la vente à Gbemba

Photo 12. Coupe de bois à Djangbétakou

37La coupe et la récolte de paille en savane est une activité traditionnelle (Figure 19). Lors de nos enquêtes, plusieurs tas ou coupeurs en activité ont été repérés. La paille est récoltée pour la confection des cases. Cette activité pose moins de problèmes pour la réserve, car la paille de savane est une ressource renouvelable abondante.
Figure 19. Coupe de paille

38La mise en culture des terres de la réserve concerne principalement sa zone périphérique. Ce phénomène traduit le déficit de sols cultivables face à la croissance naturelle de la population riveraine et aux flux réguliers d'immigrants. Aux alentours de la réserve, les pratiques culturales ont beaucoup contribué à l'érosion et à la dégradation des sols (Figure 20 et 21). Les cultures traditionnelles sur brûlis sont pratiquées par presque tous les paysans (90 %). Cette méthode culturale appauvrit les sols à cause de l'utilisation du feu pour préparer les parcelles et augmente les risques d'érosion éolienne et hydrique. En causant une baisse de la productivité agricole, ces phénomènes sont responsables de la pression foncière poussant les populations à chercher et à défricher des terres nouvelles plus fertiles pour la culture, notamment dans les zones périphériques de la réserve où s’installent en priorité les immigrants avec l’accord des communautés locales. Ces mises en culture sont plus développées dans la partie ouest de la réserve et qui débordent alors de la périphérie vers le centre de l’aire protégée (Figure 7).
Figure 20. Maraichage à Sanloaga

Figure 21. Culture de Maïs au bord de l’Oti

39Les feux de brousse (Figure 22) entraînent la perte de la matière organique végétale qui réduit la fertilité des sols et occasionne une baisse de leur productivité. Les éleveurs allument des feux en savane et dans les plaines inondées à graminées pérennes pour stimuler la reprise de la croissance et de disposer d'herbe verte de grande qualité nutritive. Lorsqu’ils sont allumés en zone sud-sahélienne, où le fourrage herbacé est formé d'espèces annuelles, ils détruisent entièrement le tapis herbacé privant ainsi le bétail de pâturage. Les feux ont des effets néfastes sur la végétation ligneuse. Les pousses annuelles fragiles meurent, compromettant ainsi la régénération de la végétation. Selon Mitja et Puig (1990) en zone de savane, le cycle de la phytomasse aérienne de la strate herbacée est liée aux feux qui la détruisent en grande partie chaque année. Les feux précoces consument les fruits et graines sur les tiges ou à la surface du sol, occasionnant une modification de la composition floristique du tapis herbacé d'une année à l'autre. En détruisant la végétation herbacée et en réduisant le recouvrement ligneux, les feux dégradent les habitats indispensables à la faune. Les populations de reptiles, oiseaux, batraciens, insectes qui dépendent des micro-milieux de la strate herbacée sont réduites. Les petits animaux comme les insectes marcheurs qui ne peuvent pas échapper aux feux sont tués. Les feux bien conduits constituent une méthode de gestion des pâturages entraînant une amélioration de la productivité et de la valeur nutritive des fourrages. Dans le cas contraire, ils pourraient compromettre l’équilibre général de l’écosystème.
Figure 22. Effets nocifs du feu sur la régénération et les peuplements d’arbres de la réserve de l’Oti-Mandouri

40On note aussi des installations illégales de campements de Peuhls dans la réserve de l’Oti-Mandouri.
41En plus des nationaux, les auteurs de la plupart des activités dans la réserve sont généralement des individus venus des pays sahéliens (les transhumants) et du Bénin voisin.
42Pour mesurer l’intensité de prélèvement des ressources végétales dans la réserve, un inventaire des souches d’individus coupés ou écorcés (prélèvement d’écorces) a été effectué et a permis de calculer les taux de prélèvements par formation et par espèces (Tableau 1).
Tableau 1. Prélèvement d’organes dans les formations végétales
|
Type de formation
|
Densité (Tiges/ha)
|
Nombre de tiges prélevées/ha
|
Taux de prélèvement (%)
|
Nombre d’espèces prélevées
|
Principales espèces prélevées
|
|
Forêt galerie
|
940
|
140
|
14 ,89
|
10
|
Pterocarpus santalinoides, Cola laurifolia,
Cynometra microphyla
|
|
Savane boisée
|
288,89
|
122,22
|
42,30
|
7
|
Pseudocedrela kotschyi, Mitragyna inermis
Daniellia oliveri
|
|
Savane arborée
|
477,78
|
166,67
|
34,88
|
17
|
Sclerocarya birrea, Prosopis africana, Sterculia setigera, Lannea spp, Terminalia spp, Anogeisus leiocarpus
|
|
Savane arbustive
|
566,67
|
144
|
25,41
|
15
|
Pteleopsis suberosa, Piliostigma thonningii, Pterocarpus erinaceus, Pericopsis laxiflora,
|
43Les Combretaceae et les Mimosaceae, indicatrices d’un climat généralement sec (Aubréville, 1950) sont dominantes et omniprésentes dans cette zone d’étude située dans la région septentrionale du nord Togo. Cela s’explique par la faiblesse des précipitations et des températures très élevées qui traduisent l’aridité du climat soudanien. La dominance numérique des Mimosaceae et Combretaceae est comparable aux résultats de Thiombiano (1996). Toutefois, l’émergence des Rubiaceae constitue une particularité qui serait liée à la grande forêt galerie le long de l’Oti. L’analyse floristique de la réserve de faune de l’Oti-Mandouri montre qu’elle renferme un potentiel ligneux non négligeable en considérant la diversité des ligneux inventoriés (116 ligneux), soit un peu plus (le double) du nombre d’espèces ligneuses recensé par Ganaba (1994) et Ouédraogo (2006) qui ont respectivement inventorié 55 et 53 espèces ligneuses au Burkina Faso.
44La réserve Oti-Mandouri est soumise à diverses formes de pression. Étant donné que la plupart des terres fertiles sont localisées à l’intérieur de la réserve, elles sont de ce fait convoitées par les agriculteurs et font ainsi l’objet de défrichements agricoles. Outre l’envahissement agricole, d’autres menaces pèsent sur la biodiversité mise en protection : la transhumance, la chasse, les coupes de bois, les cueillettes, etc. Selon Benoit (1998) in Convers et al. (2007), l’impact de la transhumance sur les ressources naturelles est essentiellement lié à la crise environnementale qui sévit dans la région ouest-africaine depuis les épisodes de grave sécheresse des années 1970 et qui se perpétue depuis avec l’augmentation de la densité démographique, en présence de populations dont le mode de vie a toujours largement reposé sur le libre accès aux ressources naturelles et à leur exploitation. Les formations naturelles sont ainsi de plus en plus fragmentées en îlots enrobés dans les paysages agraires (Hansen et al., 2000). Dans la réserve de l’Oti-Mandouri, les conflits entre forestiers et éleveurs sont fréquents à cause de la présence de troupeaux illégaux dont les impacts sur ses écosystèmes peuvent s’avérer préjudiciables (Toutain et al., 2003). Les travaux de Sarr (2010) dans le parc national des "deux balé" au Burkina Faso ont montré que la circulation du bétail crée des pistes et des zones de piétinement excessif partout, notamment autour des mares et dans les prairies attenantes. Ces différentes activités qui étaient autrefois interdites dans ladite réserve sont aujourd’hui menées par les populations riveraines à cause de l’augmentation de la population et de la paupérisation croissante. Les produits prélevés sont destinés aussi bien à l’utilisation domestique qu’à la vente. Ces activités ont été régulièrement réprimées par les agents forestiers qui arrêtent les individus en infraction, confisquent leurs matériels d’exploitation, détruisent les champs et les produits saisis ou leur infligent des amendes. Tout ceci conduit à des rapports tendus entre forestiers et les autochtones (Atutonu, 2005).
45Ces activités ont pour conséquences la raréfaction de certaines espèces végétales, la diminution de la couverture forestière, la perte d’habitats des animaux et de biodiversité, l’érosion des sols le changement des régimes hydrauliques, la perturbation climatique, etc.
46Les facteurs anthropiques perturbent fortement le fonctionnement des écosystèmes de ladite réserve engendrant ainsi la modification de la composition floristique et de la structure de la végétation (Sinsin, 1994, 2001 ; Hahn-Hadjali, 1998 ; Fournier et al., 2002). Les taux de prélèvement calculés sont très élevés par rapport à ceux calculés par Borozi (2006) ce qui montre que la réserve est soumise à de fortes pressions anthropiques, destructrices pour la végétation de la réserve. Cette situation peut être expliquée par le fait qu’il y a un manque de rigueur et de suivi dans l’application des textes, les gestionnaires ressentent des menaces liées au climat politique local. De plus, ils reconnaissent subir des pressions pour exploiter de manière excessive les ressources dans la réserve. Ainsi, de l’avis général, les pots de vin et la corruption sont fréquents, à tous les niveaux.
47Dans cette réserve, les feux de brousse sont fréquents et entraînent la destruction des habitats des animaux conduisant à la réduction du potentiel faunique disponible. Il est à noter que la disparition de certaines espèces animales peut ralentir le rythme de dissémination de certaines espèces végétales dont les fruits sont transportés par les animaux d’un milieu à un autre ou doivent passer par le tube digestif d’un animal pour faciliter la germination ; c’est le cas par exemple des éléphants.
48En effet, à cause de la croissance démographique galopante, de la pauvreté et le coût des soins médicaux élevés, les populations riveraines font beaucoup plus recours aux plantes de la réserve pour se soigner ; ce qui explique une forte pression sur les espèces végétales à usage médical.
49L’étude a permis de mettre en évidence les principales activités menées dans la réserve et leur distribution spatiale. Cette emprise sur les ressources de la réserve démontre les limites du système de gestion mis en place. En effet, le manque de moyens de déplacement et de contrôle, l’insuffisance de sensibilisation des populations riveraines et le nombre insuffisant des agents forestiers constituent autant de problèmes qui freinent la bonne gestion de cette réserve. La gestion durable de la réserve ne peut être effective que si l’implication des populations locales aux actions de conservation de la diversité biologique est active, volontaire et que les acteurs de développement qui interviennent dans le milieu travaillent en étroite collaboration avec celles-ci en vue d’identifier leurs problèmes et les intégrer dans leur programme d’action.
50Il serait donc souhaitable de mener des actions idoines pour préserver au maximum tout ce qui reste dans cette réserve, car sa proximité avec le complexe d’aires protégées appelé WAP (Parc W au Niger, parc Arly Burkina Faso et la réserve de biosphère de Pendjari au Bénin) laisse ouverte la possibilité d’un repeuplement ultérieur naturel depuis ces zones, et la possibilité de maintenir un corridor vers le parc national d’Oti-Kéran. Ceci permettra de valoriser les potentialités touristiques et par là créer des sources de revenus pour les populations locales.
51Les auteurs remercient l’IRD et l’AIRD pour leur appui financier à travers le projet RIPIECSA (Recherche Interdisciplinaire et Participative sur les Interactions entre les Écosystèmes, le Climat et les Sociétés d’Afrique de l’Ouest) pour la réalisation de ce travail. Ces remerciements vont également à l’endroit des gestionnaires et autres acteurs de terrain (personnels et populations des différents villages visités) pour leur disponibilité et leur amabilité.






































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}