DISCUSSIONThe analyses above do not paint a clear picture of the locomotor habit of Protypotherium, but they do suggest it was not highly specialized for any particular mode of locomotion. In neither PCA does Protypotherium plot near the edge of the modern mammal morphospace. Instead, it is close to the center with mammals of varying locomotor habits, implying a generalized morphology. The DFA of linear measurements indicates some aspects of Protypotherium skeletal morphology are unusual relative to the modern mammals considered, but the DFA of functional indices does not show a similar pattern. Both DFAs suggest an arboreal or semifossorial lifestyle for Protypotherium, with the latter having a higher conditional probability. The functional indices themselves do not suggest a single locomotor style. BI of Protypotherium is < 100, similar to hyraxes and some arboreal and semifossorial mammals; CI of Protypotherium is > 100, close to values seen in cursorial canids (Appendix). Few mammals show as great a discrepancy between the relative lengths of fore- and hind-limbs and this could be a phylogenetic characteristic of notoungulates. The relatively high EI of Protypotherium resembles that of semifossorial and arboreal mammals, but MFI is higher than one would expect for an arboreal mammal (thus favoring semifossoriality). Qualitatively, the limb bones of Protypotherium also show a mix of characters; the proximal and distal elements share features with generalized and cursorial mammals, whereas the intermediate elements more closely resemble semifossorial ones. So was Protypotherium a generalist, or did it have a more specialized locomotor habit? Because all notoungulates are extinct, it is impossible to unequivocally 'calibrate' morphology and behavior to determine the locomotor habit of a notoungulate with a generalized skeleton. As an analogy, even though Dasyprocta is a cursorial rodent, one might not make such an inference based solely on comparisons with non-rodents. DFAs of our dataset bear this out; Dasyprocta was sometimes classified as generalized when other rodents were excluded. Clearly inferences about extinct taxa are biased to some degree by the extant animals available for comparison. This holds true for all extinct species, but the problem is exacerbated in mammals like notoungulates (and litopterns, uintatheres, palaeanodonts, etc.) in which the entire order is extinct (as opposed to just the suborder or family). That being said, it is unlikely that the skeleton of a committed runner (or digger or climber) would lack any characteristics suggestive of such habits. It is therefore unlikely that Protypotherium spent all of its time either running, digging, or climbing. On the other hand, it is just as unlikely that it spent an equal proportion of its time in each of those pursuits; interpreting it merely as a 'generalist' is neither very enlightening nor very satisfying. The overall attributes of the Protypotherium postcranial skeleton suggest a locomotor habit tending toward cursoriality; moment arms for muscles mostly favor speed over force and articulations tend more to restrict movement than to increase mobility. The digits are reduced – most compatible with a terrestrial/cursorial habit – and the unguals show no specializations for excavation or climbing. Qualitatively, Protypotherium is better designed for cursoriality than anything else, though it certainly is not proportioned like a cursor nor highly designed for that mode of locomotion. A skeleton more adapted for force than speed is suggested by the DFAs, but this may be a phylogenetic effect rather than evidence for a particular mode of locomotion. As noted above, Protypotherium is being judged against other clades of mammals; such comparisons may not discriminate a slightly more arboreal (scansorial) notoungulate from a slightly more cursorial one, especially if the ancestral locomotor habit is fossoriality (see discussion in Shockey et al. 2007). A notoungulate tending toward cursoriality may look like a generalized or semifossorial carnivoran (or rodent) merely because it started out further away from that end of the locomotor spectrum and still bears features characteristic of its ancestry. Moreover, although most of the important moment arms of the appendicular skeleton were used as variables in the quantitative analyses, these only describe a small proportion of overall skeletal morphology. For example, the relative lengths of the ulna and olecranon are functionally important and vary predictably with locomotor habit (e.g., Vizcaíno and Milne 2002), but neither of these metrics describes the shape of the olecranon, the morphology of the semilunar notch, or how the radial head articulates with the ulna – additional attributes of functional importance (e.g., Rose 1990, 2001). The subset of postcranial measurements used might not be providing the same picture of locomotor habit as that of a larger, more comprehensive dataset.
Indirect support for cursorial tendencies in Protypotherium comes from two other Miocene interatheriids: Miocochilius and Interatherium. Miocochilius is best known from the middle Miocene of Colombia (Stirton 1953;
Kay et al. 1997). It is likely the sister-taxon to Protypotherium (Reguero et al. 2003) and although the two genera are similar craniodentally (Hitz et al. 2000;
Reguero et al. 2003;
Croft 2007) they differ postcranially; most conspicuously, Miocochilius bears only two functional digits on the fore- and hind limbs (Stirton 1953;
Figure 4 and
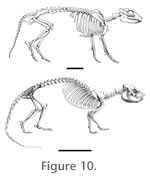
Figure 10). I |
|