Results
Each symbol (grey circle or colored sphere) on scatter plots in figures of this paper represents a composite OTU, which is the set of measurements for nine morphological characters from a single fenestellid colony. When viewing and interpreting the PCA scatter plots, the reader should keep in mind that: 1) each symbol represents an "observation" from a colony = nine measurements; 2) each color represents a bryozoan systematist's generic identification for the specimen from which the data were collected; 3) each group of symbols representing a genus consists of a variable number of species, colonies and composite observations per colony; and 4) for the most part, the original and traditional generic diagnoses for these taxa did not include any of the nine characters used in these plots (i.e., these plots represent relationships among characteristics typically not used to define the taxonomic concept of any of the genera).
The first results section characterizes each Principal Component Axis, i.e., which characteristics are most important for the axis and which regions of the plots will reflect which characters. This section also describes whether the axes differentiate variation among genera (have taxonomic value) or variation within genera (variation among and within species and individuals).
 Details are reserved for the second results section, which summarizes the distribution of genera within the defined morphospace.
Details are reserved for the second results section, which summarizes the distribution of genera within the defined morphospace.
Among Genera vs. Within Genera Variation. The distribution of specimens expressed on Principal Component Axes one through five (PCA-1 to PCA-5) primarily reflect variation among genera and collectively account for 82.2% of the observed variation (Table 3).
 The relationship between the positions of data points (colony-level observations) and the character states for morphological features with which the axes are most correlated is represented in
Figure 3 for PCA-1 and PCA-3. These same data, plotted on PCA-1, PCA-2, and PCA-3, are color coded to represent generic assignment in
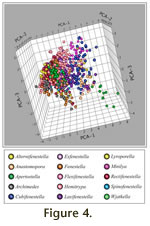
Figure 4, which is linked to an animation of the axes in rotation (Animation 1). All animations in this paper can be accessed either directly by hyperlinks from the text, or via the
Animation Menu. The relationship between the positions of data points and the character states for morphological features with which the axes are most correlated for PCA-2 and PCA-3 is represented in
Figure 5.
The relationship between the positions of data points (colony-level observations) and the character states for morphological features with which the axes are most correlated is represented in
Figure 3 for PCA-1 and PCA-3. These same data, plotted on PCA-1, PCA-2, and PCA-3, are color coded to represent generic assignment in
Figure 4, which is linked to an animation of the axes in rotation (Animation 1). All animations in this paper can be accessed either directly by hyperlinks from the text, or via the
Animation Menu. The relationship between the positions of data points and the character states for morphological features with which the axes are most correlated for PCA-2 and PCA-3 is represented in
Figure 5.
 These same data, plotted on PCA-1, PCA-2, and PCA-3, are color coded to represent generic assignment in
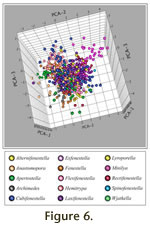
Figure 6, which is linked to an animation of the axes in rotation (Animation 1). The relationship between the positions of data points and the character states for morphological features with which the axes are most correlated for PCA-4 and PCA-5 is represented in
Figure 7.
These same data, plotted on PCA-1, PCA-2, and PCA-3, are color coded to represent generic assignment in
Figure 6, which is linked to an animation of the axes in rotation (Animation 1). The relationship between the positions of data points and the character states for morphological features with which the axes are most correlated for PCA-4 and PCA-5 is represented in
Figure 7.
 These same data, plotted on PCA-3, PCA-4, and PCA-5, are color coded to represent generic assignment in
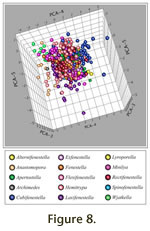
Figure 8, which is linked to an animation of the axes in rotation (Animation 2).
These same data, plotted on PCA-3, PCA-4, and PCA-5, are color coded to represent generic assignment in
Figure 8, which is linked to an animation of the axes in rotation (Animation 2).
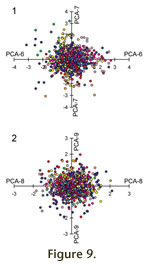
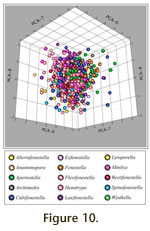
Principal Component Axes six through nine (PCA-6 to PCA-9) primarily reflect variation within genera and collectively account for 17.8% of the observed variation (Table 3). Data points (colony-level observations color coded to represent generic assignment) are plotted on PCA-6 and PCA-7, and on PCA-8 and PCA-9 in
Figure 9. These same data are plotted on PCA-6, PCA-7, and PCA-8 in
Figure 10, which is also linked to an animation of the axes in rotation (Animation 3).
PCA-1: Size.
 Principal Component Axis One represents overall size for all characters (Table 4, column 1) and provides differentiation among most genera (Table 5,
Figure 3). All characters are about equally weighted, with emphasis on fenestrule size (FL and FW), slightly less weight on Aperture spacing Laterally across Branch (ALB) and node spacing and size (NS, ND), and the least on other characters.
Principal Component Axis One represents overall size for all characters (Table 4, column 1) and provides differentiation among most genera (Table 5,
Figure 3). All characters are about equally weighted, with emphasis on fenestrule size (FL and FW), slightly less weight on Aperture spacing Laterally across Branch (ALB) and node spacing and size (NS, ND), and the least on other characters.
In Figure 3 and
Figure 4, specimens with large values for all characters plot high on PCA-1 (positive = right on
Figure 3) and those with the
collectively smallest values for all characters plot low (negative = left) on
PCA-1.
 There
is a finite size limit on the negative part of PCA-1 (absolute smallest size in
reality and observed). As a result, there is more room to expand in size and
specialize in the positive direction of PCA-1 (minimum value is -3.5, whereas
the maximum value is 8.0) (Figure 3 and
Figure 4). Values are negatively skewed on PCA-1 with a median of -0.289. The size trend is evident when the data cloud is viewed in three dimensions PCA-1 vs. PCA-2, vs. PCA-3 (Figure
4). By definition (all characters having approximately equal weight on PCA-1) large values are excluded from each of the remaining axes when scores are low on PCA-1. This autocorrelation results in a conical shape of the overall cloud (narrowing to the left on
Figure 3 and
Figure 4 and when PCA-1 is included in any scatter plot.
There
is a finite size limit on the negative part of PCA-1 (absolute smallest size in
reality and observed). As a result, there is more room to expand in size and
specialize in the positive direction of PCA-1 (minimum value is -3.5, whereas
the maximum value is 8.0) (Figure 3 and
Figure 4). Values are negatively skewed on PCA-1 with a median of -0.289. The size trend is evident when the data cloud is viewed in three dimensions PCA-1 vs. PCA-2, vs. PCA-3 (Figure
4). By definition (all characters having approximately equal weight on PCA-1) large values are excluded from each of the remaining axes when scores are low on PCA-1. This autocorrelation results in a conical shape of the overall cloud (narrowing to the left on
Figure 3 and
Figure 4 and when PCA-1 is included in any scatter plot.
PCA-2:
 Aperture Size vs. Spacing, Plus Dissepiment Width. PCA axis two represents an inverse relationship between the size of an aperture (AD) vs. aperture spacing along the branch (AAB) and provides differentiation among several genera (Table 5,
Figure 5). Thus, small apertures have greater spacing between them (Table 5). Specimens with large values for Aperture Diameter (AD) plot low on PCA-2 (to the left on
Figure 5) and those with large aperture spacing along branch (AAB) plot high on PCA-2 (to the right on
Figure 5). Axis two also displays a positive relationship between the size of apertures (AD) and Dissepiment Width (DW), e.g., wide dissepiments are associated with more closely spaced apertures along branch (Table 5). Several genera are differentiated on PCA-2, especially those that plot relatively high on PCA-1. Otherwise, the specimen cloud is centered near zero on PCA-2 (Figure 5).
Aperture Size vs. Spacing, Plus Dissepiment Width. PCA axis two represents an inverse relationship between the size of an aperture (AD) vs. aperture spacing along the branch (AAB) and provides differentiation among several genera (Table 5,
Figure 5). Thus, small apertures have greater spacing between them (Table 5). Specimens with large values for Aperture Diameter (AD) plot low on PCA-2 (to the left on
Figure 5) and those with large aperture spacing along branch (AAB) plot high on PCA-2 (to the right on
Figure 5). Axis two also displays a positive relationship between the size of apertures (AD) and Dissepiment Width (DW), e.g., wide dissepiments are associated with more closely spaced apertures along branch (Table 5). Several genera are differentiated on PCA-2, especially those that plot relatively high on PCA-1. Otherwise, the specimen cloud is centered near zero on PCA-2 (Figure 5).
 PCA-3: Fenestrule Size vs. Aperture Size and Spacing. Axis three represents an inverse relationship between the size (openness) of fenestrules and the size and spacing of apertures and provides differentiation among several genera (Table 5,
Figure 5). Specimens with larger, more open fenestrules and small, closely spaced apertures plot low on PCA-3 (to the bottom of
Figure 5), whereas those with smaller fenestrules and larger, more distantly spaced apertures plot high (to the top on
Figure 5). Principal Component axis three also reflects an inverse relationship between node spacing vs. size (NS vs. ND). Specimens with larger diameter nodes plot higher on PCA-3 (to the top on
Figure 5), whereas those with a greater spacing between nodes plot lower on PCA-3 (to the bottom on
Figure 5). The distribution of samples on PCA-3 can also be evaluated on
Figure 6.
PCA-3: Fenestrule Size vs. Aperture Size and Spacing. Axis three represents an inverse relationship between the size (openness) of fenestrules and the size and spacing of apertures and provides differentiation among several genera (Table 5,
Figure 5). Specimens with larger, more open fenestrules and small, closely spaced apertures plot low on PCA-3 (to the bottom of
Figure 5), whereas those with smaller fenestrules and larger, more distantly spaced apertures plot high (to the top on
Figure 5). Principal Component axis three also reflects an inverse relationship between node spacing vs. size (NS vs. ND). Specimens with larger diameter nodes plot higher on PCA-3 (to the top on
Figure 5), whereas those with a greater spacing between nodes plot lower on PCA-3 (to the bottom on
Figure 5). The distribution of samples on PCA-3 can also be evaluated on
Figure 6.
PCA-4: Branch and Dissepiment Width vs. Node Size and Spacing and Aperture Size. Axis four represents an inverse relationship between the combined size (width) of branches and dissepiments (= robustness) relative to the size and spacing of nodes and aperture size (diameter) (Table 4). PCA-4 provides some differentiation among several genera (Table 5,
Figure 7). Specimens with wider branches and dissepiments and smaller more closely spaced nodes plot low (to the left on
Figure 7), whereas those with more narrow branches and dissepiments and larger more distantly spaced nodes plot high (to the right on
Figure 7). The distribution of samples on PCA-4 can also be evaluated on
Figure 8.
Most taxa cluster near the center of PCA-4 (zero on
Figure 7), but some generic groups do differentiate along PCA-4, so therefore combined characters BW, DW, ND, NS and AD have significance for differentiating among some fenestellid genera.
PCA-5: Dissepiment Width, Aperture Spacing, and Node Diameter vs. Aperture Size and Spacing. Axis five does not represent a simple relationship among characters with obvious biological links (Table 5). Specimens with larger apertures (AD) and apertures more distantly spaced laterally across a branch (ALB) plot high on PCA-5 (to top on
Figure 7). Specimens with wide dissepiments, large nodes, and close aperture spacing along branches plot low on PCA-5 (to bottom on
Figure 7). The distribution of samples on PCA-5 can also be evaluated on
Figure 8. Although specimens are centered on a mean of zero for PCA-5 (Figure 7), considerable variation exists toward negative values on the axis (bottom on
Figure 7). Some variation among genera is visible along PCA-5.
PCA-6: Branch Width and Node Size/Spacing vs. Dissepiments Width and Aperture Size/Spacing. Axis six reflects an inverse relationship between wider branches with larger, more distantly spaced nodes as compared to narrower dissepiments and with smaller more closely spaced apertures (Table 6). Specimens with wide dissepiments (DW) and larger more distantly spaced apertures (ALB and AD), plot high on PCA-6 (to right on
Figure 9.1). Specimens with wide branches and large distantly spaced nodes plot low on PCA-6 (to left on
Figure 9.1). Although specimens are centered on a mean of zero for PCA-6, values are scattered from -3.0 to +3.0 on Axis six (Figure 9.1). The distribution of samples on PCA-6 can also be viewed on
Figure 10.
PCA-7: Fenestrule Width and Node Diameter vs. Node and Aperture Spacing. Axis seven reflects an inverse relationship between wider fenestrules with larger nodes as compared to more closely spaced nodes and apertures (Table 6). Specimens with wide fenestrules (FW) and large nodes (ND), plot low on PCA-7 (to bottom of
Figure 9.1). Specimens with more distantly spaced nodes (NS) and apertures along branch (AAB) plot high on PCA-7 (to top on
Figure 9.1). Specimens are centered on a mean of zero for PCA-7, but values are scattered from -3.0 to +3.0 on Axis seven (Figure 9.1). The distribution of samples on PCA-7 can also be viewed on
Figure 10.
PCA-8: Fenestrule Width and Aperture Size/Spacing vs. Node Diameter, Fenestrule Length and Aperture Spacing. Axis eight reflects an inverse relationship between larger apertures more distantly spaced along branches and wider fenestrules as compared to more distantly spaced apertures laterally across branches, large nodes, and long fenestrules. Specimens with wide fenestrules (FW) and large apertures (AD) that are distantly spaced along branch (AAB), plot low on PCA-8 (to left of
Figure 9.2). Specimens with large nodes (ND), long fenestrules (FL), and distantly spaced apertures laterally (ALB) plot high on PCA-8 (to right on
Figure 9.2). Specimens are centered on a mean of zero for PCA-8, but values are restricted to only -2.0 to +2.0 on Axis eight (Figure 9.2). The distribution of samples on PCA-8 can also be viewed on
Figure 10.
PCA-9: Node and Aperture Spacing and Fenestrule Width vs. Fenestrule Length and Aperture Diameter. Axis nine reflects a complex inverse relationship between more distantly spaced nodes and apertures as compared to narrower fenestrules with large apertures. Specimens with distantly spaced nodes (NS) and lateral spacing of apertures (ALB) and wide fenestrules (FW), plot low on PCA-9 (to bottom of
Figure 9.2). Specimens with long fenestrules and large apertures plot high on PCA-9 (to top on
Figure 9.2).
Distribution of Genera in Morphospace
Fenestellid genera are compared here in a morphospace defined by the first three principal components, which account for 64.9% of variance and represent 1) overall size (small vs. large); 2)
 aperture spacing along branch vs. aperture size and dissepiment width (AD vs. AD, DW); and 3) node size, aperture spacing along branch and aperture diameter vs. fenestrule size and node spacing (ND, AAB vs. FL, FW, NS). Although axes four and five also represent among genera variance, trends are less general and are only reported in cases of relevance. In the following descriptions, "clouds" or "clusters" of OTUs are described in morphospace, but the values provided are intended to direct the reader's eye, not to provide absolute ranges for observed values
(Table 7).
aperture spacing along branch vs. aperture size and dissepiment width (AD vs. AD, DW); and 3) node size, aperture spacing along branch and aperture diameter vs. fenestrule size and node spacing (ND, AAB vs. FL, FW, NS). Although axes four and five also represent among genera variance, trends are less general and are only reported in cases of relevance. In the following descriptions, "clouds" or "clusters" of OTUs are described in morphospace, but the values provided are intended to direct the reader's eye, not to provide absolute ranges for observed values
(Table 7).
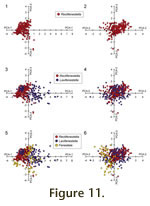
Data points (colony-level observations), color coded to represent the genera Rectifenestella, Laxifenestella, and Fenestella, are plotted on PCA-1 and PCA-3, and on PCA-2 and PCA-3 in
Figure 11.
 These same data are plotted on PCA-1, PCA-2, and PCA-3 in
Figure 12 with all data points from the study included as small black spheres for reference. Genera are highlighted in groups as Rectifenestella (Figure 12.1, axes animated in rotation in
Animation 4); Laxifenestella and Rectifenestella (Figure 12.2, axes animated in rotation in
Animation 5); Fenestella, Laxifenestella, and Rectifenestella (Figure 12.3, axes animated in rotation in
Animation 6); and Hemitrypa, Rectifenestella, and Lyroporella (Figure 12.4, axes animated in rotation in
Animation 7).
These same data are plotted on PCA-1, PCA-2, and PCA-3 in
Figure 12 with all data points from the study included as small black spheres for reference. Genera are highlighted in groups as Rectifenestella (Figure 12.1, axes animated in rotation in
Animation 4); Laxifenestella and Rectifenestella (Figure 12.2, axes animated in rotation in
Animation 5); Fenestella, Laxifenestella, and Rectifenestella (Figure 12.3, axes animated in rotation in
Animation 6); and Hemitrypa, Rectifenestella, and Lyroporella (Figure 12.4, axes animated in rotation in
Animation 7).
 Data points, color coded to represent each genus in this study, are plotted in several combinations on PCA-4 and PCA-5 in
Figure 13. Combinations of genera included in
Figure 13 are 1) Rectifenestella, Laxifenestella and Fenestella; 2) Rectifenestella, Hemitrypa, and Lyroporella; 3) Rectifenestella, Fenestella, and Archimedes; 4) Rectifenestella, Cubifenestella, and Apertostella; 5) Alternifenestella, Minilya, and Spinofenestella; and 6) Exfenestella, Flexifenestella, Anastomopora, and Wjaktella.
Data points, color coded to represent each genus in this study, are plotted in several combinations on PCA-4 and PCA-5 in
Figure 13. Combinations of genera included in
Figure 13 are 1) Rectifenestella, Laxifenestella and Fenestella; 2) Rectifenestella, Hemitrypa, and Lyroporella; 3) Rectifenestella, Fenestella, and Archimedes; 4) Rectifenestella, Cubifenestella, and Apertostella; 5) Alternifenestella, Minilya, and Spinofenestella; and 6) Exfenestella, Flexifenestella, Anastomopora, and Wjaktella.
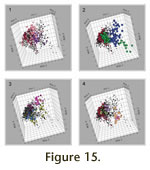
 Data points, color coded to represent the genera in groups of Archimedes, Fenestella, and Rectifenestella, and Cubifenestella, Apertostella, and Rectifenestella, are plotted on PCA-1 and PCA-3, and on PCA-2 and PCA-3 in
Figure 14. The data for these two groups of genera are plotted on PCA-1, PCA-2, and PCA-3 in
Figure 15. The group that highlights Archimedes (Figure 15.1) has axes animated in rotation in
Animation 8). The group that highlights Cubifenestella and Apertostella (Figure 15.2) has axes animated in rotation in
Animation 9.
Data points, color coded to represent the genera in groups of Archimedes, Fenestella, and Rectifenestella, and Cubifenestella, Apertostella, and Rectifenestella, are plotted on PCA-1 and PCA-3, and on PCA-2 and PCA-3 in
Figure 14. The data for these two groups of genera are plotted on PCA-1, PCA-2, and PCA-3 in
Figure 15. The group that highlights Archimedes (Figure 15.1) has axes animated in rotation in
Animation 8). The group that highlights Cubifenestella and Apertostella (Figure 15.2) has axes animated in rotation in
Animation 9.
 Data points, color coded to represent the genera, grouped as 1) Alternifenestella, Minilya, and Spinofenestella and 2) Exfenestella, Flexifenestella, Anastomopora, and Wjaktella, are plotted on PCA-1 and PCA-3 and on PCA-2 and PCA-3 in
Figure 16. The first group of data (Figure 16.1) has axes animated in rotation in
Animation 10, and the second group (Figure 15.2) has axes animated in rotation in
Animation 11.
Data points, color coded to represent the genera, grouped as 1) Alternifenestella, Minilya, and Spinofenestella and 2) Exfenestella, Flexifenestella, Anastomopora, and Wjaktella, are plotted on PCA-1 and PCA-3 and on PCA-2 and PCA-3 in
Figure 16. The first group of data (Figure 16.1) has axes animated in rotation in
Animation 10, and the second group (Figure 15.2) has axes animated in rotation in
Animation 11.
 Rectifenestella. The genus Rectifenestella
Morozova, 1974 was originally diagnosed as a fenestellid having straight branches and dissepiments, pentagonal cross sections of endozonal zooecial chambers, and a single series of keel nodes (Morozova 1974). It was chosen as the fenestellid genus for standard of comparison here because of its basic form, large number of observations (243), and number of nominal species (seven) widely scattered both stratigraphically and geographically (Appendix). The distribution of Rectifenestella in morphospace can be summarized as follows.
Rectifenestella. The genus Rectifenestella
Morozova, 1974 was originally diagnosed as a fenestellid having straight branches and dissepiments, pentagonal cross sections of endozonal zooecial chambers, and a single series of keel nodes (Morozova 1974). It was chosen as the fenestellid genus for standard of comparison here because of its basic form, large number of observations (243), and number of nominal species (seven) widely scattered both stratigraphically and geographically (Appendix). The distribution of Rectifenestella in morphospace can be summarized as follows.
1. On Figure 11.1 there is a highly coherent cluster to the left side almost centered on PCA-3 (PCA-1 values from -2.5 to 0.0 and PCA-3 values from -2. to 2.0). The cluster is bounded on the right by a sharp boundary (PCA-1 0.0) and tapers to the left converging on PCA-1 and PCA-3 values of -3.0 and -0.5.
2. On
Figure 11.2 there is a coherent cluster centered to the right of the origin (0.5, 0.5) with a range of PCA-2 values from -2.0 to 2.0 and PCA-3 values from -2.0 to 2.0).
3. On
Figure 13.1 there is a single, coherent cluster centered on the origin (PCA-4 values from -1.0 to 1.0 and PCA-5 values from -1.5 to 1.5).
Laxifenestella. The genus Laxifenestella
Morozova, 1974 was originally diagnosed as a fenestellid having straight to slightly sinuous branches, moderately broad dissepiments, well-developed superior hemisepta, tetragonal to pentagonal cross sections of endozonal zooecial chambers, and small, closely spaced keel nodes (Morozova 1974). It is represented here by a large number of observations (117) and six nominal species with broad geographic and stratigraphic distribution. The distribution of Laxifenestella in morphospace falls into two discrete subclusters and can be summarized as follows.
1. On Figure 11.3 there is a tight subcluster in the upper left quadrant (PCA-1 values from -2.0 to -1.0 and PCA-3 values from 0.5 to 1.5). A second, more diffuse subcluster is found in the right region centered on PCA-3 (PCA-1 values from 0.0 to 4.0 and PCA-3 -2.0 to 1.5). The first subcluster (all OTUs assigned to Laxifenestella serratula (Ulrich) by
Snyder (1991)) coincides with observations from Fenestella and Rectifenestella.
2. On
Figure 11.4 there is a single, diffuse cluster centered on the origin (PCA-2 values from -2.0 to 2.0 and PCA-3 values from -2.0 to 1.5).
3. On
Figure 13.1 there is a single diffuse cluster centered on the lower left quadrant (PCA-4 values from -2.0 to 1.5 and PCA-5 values from -2.5 to 1.5).
Fenestella. The genus Fenestella
Lonsdale, 1839, was the second fenestrate genus named, for zoaria "composed of branches which unite by growth and form a cup," characterized by "one row" of zooecia on branches and none on branch interconnections. It is the most well known and misapplied fenestellid genus name. It is represented here by 60 observations from five nominal species. The distribution of Fenestella in morphospace falls into two discrete subclusters and can be summarized as follows.
1. On Figure 11.5 there is a tight subcluster in the lower left quadrant (PCA-1 values from -3.0 to -2.0 and PCA-3 values from -2.0 to -1.0). The first subcluster is comprised entirely of OTUs from specimens assigned to Fenestella sp. 1 in
Ernst and Schroeder (2007). A second, more diffuse subcluster is found in the central region (PCA-1 values from -0.5 to 1.5 and PCA-3 -3.0 to 2.0). The second subcluster forms a relatively tight disk-shaped cloud, which falls in a zone between Rectifenestella and Laxifenestella.
2. On
Figure 11.6 there is a single, diffuse cluster to the left side (PCA-2 values from -2.0 to 0.0 and PCA-3 values from -3.0 to 2.0). Unlike observations for the previous two genera, Fenestella is restricted to a negative distribution on PCA-2.
3. On
Figure 13.1 there is a single cluster centered on the origin (PCA-4 values from -1.5 to 1.0 and PCA-5 values from -1.0 to 1.5).
Hemitrypa. The genus Hemitrypa
Phillips, 1841 was named for fenestrate bryozoans with a fine-textured side that proved to be a finely reticulate superstructure with each opening in the superstructure centered over a single zooecial aperture. It is represented here by 225 observations from nine nominal species. In morphospace (Figure 14.1–14.2,
Figure 12.4), Hemitrypa observations form a relatively cohesive cloud that has a complex shape. The distribution of Hemitrypa in morphospace can be summarized as follows.
1. On Figure 14.1 there is a coherent, conical cluster centered on the origin, with a pronounced taper toward the lower left quadrant converging on values of PCA-1 and PCA-3 of -3.0 and -1.0. The range of values on PCA-1 is from -3.0 to 4.0 and PCA-3 is -1.0 to 3.0. There is a horizontal boundary at PCA-3 of -1.0. The overall size of measured characters ranges from some of the smallest observations (Figure 14.1, value of -3.5 on PCA-1) to mid-sized including the range for Rectifenestella, Fenestella, and Laxifenestella. As the overall size of observed Hemitrypa increases, values extend preferentially into the upper right quadrant of
Figure 14.1.
2. On
Figure 14.2 there is a coherent cloud extending from the center to the upper left quadrant, with PCA-2 values from -2.0 to 1.0 and PCA-3 values from -1.0 to 3.0.
3. On
Figure 13.2 there is a coherent, somewhat diffuse cloud centered on the origin with PCA-4 values from -3.0 to 2.0 and PCA-5 values from -3.0 to 1.5.
Archimedes. The genus Archimedes
Owen, 1838 was the earliest Paleozoic fenestrate genus established, characterized and named on the basis of its helical support structure. It is represented here by 60 observations from five nominal species. The distribution of Archimedes in morphospace can be summarized as follows.
1. On Figure 14.3 there is a coherent cluster to the left side almost centered on PCA-3 (PCA-1 values from -2.5 to 1 and PCA-3 values from -3.0 to 1.5). The cluster is bounded on the right by an inclined boundary (PCA-1 0.5 to 1.0) and tapers to the left converging on PCA-1 and PCA-3 values of -2.0 and -1.0. Archimedes occupies a range of morphospace similar to Rectifenestella and Fenestella.
2. On
Figure 14.4 there is a coherent cluster located largely to the left of the origin (-1.0, -0.5) with a range of PCA-2 values from -2.0 to 0.5 and PCA-3 values from -3.0 to 1.5). As the overall size of Archimedes features increase, the observations are largely restricted to the negative regions of PCA-2 (to the left on
Figure 14.4).
3. On Figure 13.3 there is a single diffuse cluster centered on the origin (PCA-4 values from -1.5 to 1.5 and PCA-5 values from -0.5 to 1.5).
Cubifenestella. The genus Cubifenestella
Snyder, 1991 was originally diagnosed as a fenestellid having robust zoarial meshwork and several internal characters including absence of hemisepta, intermediate chamber size, and cubic to irregularly rectangular endozonal zooecial chambers (Snyder 1991). It is represented here by 72 observations from three nominal species. The distribution of Cubifenestella in morphospace can be summarized as follows.
1. On Figure 14.5 there is a diffuse but coherent cluster in the right side almost centered on PCA-3 (PCA-1 values from 1.5 to 6 and PCA-3 values from -2.5 to 3.0).
2. On
Figure 14.6 there is a diffuse cluster centered on the origin (PCA-2 values from -2.0 to 2.5 and PCA-3 values from -2.5 to 3.0).
3. On
Figure 13.4 there is a single, tight cluster to the right, centered on PCA-5 (PCA-4 values from -1.0 to 3.0 and PCA-5 values from -1.5 to 1.5).
Apertostella. The genus Apertostella
Snyder, 1991 was originally diagnosed as a fenestellid having delicate to robust meshwork characteristics and a complex of internal characteristics including intermediate endozonal chamber and zooecial aperture size, no hemisepta, and rectangular box shaped endozonal zooecial chambers (Snyder 1991). The genus as represented by the specimens at hand does not form a coherent cluster. The distribution of Apertostella in morphospace can be summarized as follows.
1. On Figure 14.5 there is a tight subcluster centered on the origin (PCA-1 values from -1.0 to 1.0 and PCA-3 values from -1.0 to 1.5). This subcluster occupies morphospace shared by Fenestella. A second, more diffuse subcluster is found in the lower right quadrant (PCA-1 values from 4.0 to 8.0 and PCA-3 values from -3.0 to -1.5). The second subcluster consists of OTUs entirely from specimens assigned to the species Apertostella venusta
Snyder, 1991. The second subcluster of Apertostella occupies morphospace independent of any other fenestellid genus, but adjacent to Cubifenestella.
2. On
Figure 14.6 there is a single, diffuse cluster to the left (PCA-2 values from -3.5 to 1.0 and PCA-3 values from -3.0 to 1.5).
3. On
Figure 13.4 there is a single diffuse cluster slightly offset into the upper right quadrant (PCA-4 values from -1.0 to 2.0 and PCA-5 values from -1.0 to 1.5).
Data points, color coded to represent the genera in groups of 1) Alternifenestella, Minilya, and Spinofenestella; and 2) Exfenestella, Flexifenestella, Anastomopora and Wjatkella are plotted on PCA-1 and PCA-3 and on PCA-2 and PCA-3 in
Figure 16. These same data are plotted on PCA-1, PCA-2 and PCA-3 in
Figure 15, with all data points in the study included for reference as small black spheres. Frames of
Figure 15 are linked to animations of the axes in rotation.
Minilya. The genus Minilya
Crockford, 1944 was originally diagnosed as a fenestellid having small keel nodes in two alternating rows and having sub-triangular cross sections of endozonal zooecial chambers. It is represented here by 24 observations from two nominal species. The distribution of Minilya in morphospace can be summarized as follows.
1. On Figure 16.1 there is a diffuse, elongate cloud extending into the upper right quadrant (PCA-1 values from -1.0 to 5.0 and PCA-3 values from -0.5 to 2.5).
2. On
Figure 16.2 there is a diffuse, elongate cloud in the upper right quadrant (PCA-2 values from 0.0 to 4.0 and PCA-3 values from -0.5 to 2.5).
3. On
Figure 13.5 there is a coherent, diffuse cluster inclined from the upper left to lower right quadrants (PCA-4 values from -2.5 to 2.0 and PCA-5 values from -2.5 to 2.0).
Spinofenestella. The genus Spinofenestella
Termier and Termier, 1971, was originally diagnosed as a fenestellid having a regular zoarial meshwork of short fenestrules, with a zooecial aperture at the base of each dissepiment and a single row of keel nodes (Termier and Termier 1971). It is represented by 38 observations from four nominal species. The distribution of Spinofenestella in morphospace can be summarized as follows.
1. On Figure 16.1 there is single cloud centered in the lower left quadrant (PCA-1 values from -2.0 to 1.5 and PCA-3 values from -2.0 to 0.5). Observations largely overlap the morphospace occupied by a subcluster of Alternifenestella.
2. On
Figure 16.2 there is a coherent cloud centered in the lower right quadrant (PCA-2 values from -0.5 to 1.0 and PCA-3 values from -2.0 to 0.5).
3. On
Figure 13.5 there is a single, coherent cloud in the upper central region (PCA-4 values from -1.0 to 1.0 and PCA-5 values from -1.0 to 2.0).
Alternifenestella. The genus Alternifenestella
Termier and Termier, 1971 was not originally diagnosed, being valid only because a type species was designated for it (Termier and Termier 1971).
Morozova (1974) diagnosed it as a fenestellid having thin straight branches and dissepiments, triangular to trapezoidal cross sections of endozonal zooecial chambers, frequently budded in a single row, and with a single series of keel nodes. It is represented here by 50 observations from five nominal species. The distribution of Alternifenestella in morphospace is also somewhat enigmatic and can be summarized as follows.
1. On Figure 16.1 there is a tight subcluster in the lower left quadrant (PCA-1 values from -2.0 to 1.0 and PCA-3 values from -2.0 to 0.0). A second, more diffuse subcluster is found in the right region centered on PCA-1 (PCA-1 values from 3.0 to 5.0 and PCA-3 -2.0 to 2.0). The first subcluster coincides with observations from the genus Spinofenestella.
2. On
Figure 16.2 there is a single, tight cloud in the lower right quadrant (PCA-2 values from 0.0 to 1.0 and PCA-3 values from -2.5 to 1.0), which coincides with observations from the genus Spinofenestella.
3. On
Figure 13.5 there is a single, tight cloud in upper region centered on PCA-4 (PCA-4 values from -1.0 to 1.0 and PCA-5 values from 0.0 to 1.5). This cloud coincides with observations from the genus Spinofenestella.
Other Genera. Five additional genera are represented by single species. Among these the species of Exfenestella
Morozova, 1974 and Wjatkella
Morozova, 1970 are centrally placed within the total cloud, with the centroid of their respective clouds plotting near zero on PCA-1, PCA-2, and PCA-3 (Figure 16.3-16.4). These two genera therefore occupy portions of morphospace shared with Rectifenestella, Fenestella, and Laxifenestella (Figure 16.3–16.4,
Figure 15.4). Observations of Flexifenestella
Morozova 1974 form a tight cloud in the lower right quadrant of
Figure 16.3 and plot slightly positive on PCA-2 (Figure 16.4), and negative on PCA-3 (Figure 16.3-16.4).
The multiserial genus Anastomopora
Simpson, 1897 forms a diffuse but coherent cloud centered on the right central region of
Figure 16.3 and left central region of
Figure 16.4 in the range of morphospace occupied by Cubifenestella and in part by Apertostella.
The genus Lyroporella
Simpson, 1895 is a distinctive genus characterized by transversely arched ("lyre-shaped") zoaria that are weighted by thick skeletal deposits along the proximolateral margins. It was distinguished from polyporid lyre-shaped species with multiple zooecial rows by having only two rows of zooecia that only increase to three rows proximal to branch bifurcations. Data here are restricted to 12 observations from a single nominal species. In morphospace (Figure 14.1-14.2,
Figure 12.4), Lyroporella observations plot in a compact cloud in the upper left quadrant of
Figure 14.1-14.2, in a portion of the morphospace shared by Rectifenestella and Hemitrypa.
Relationships Among Morphometric Characters
The covariance between each pair of characters is highly significant (Table 8), except for AD-ABB, which is marginally significant. Several groups of characters show a greater degree of linkage (Table 8, highlighted by color).
1. FL-FW-ALB-NS: fenestrule size (length and width), Aperture spacing Laterally across Branch, and Node Spacing display the highest pair-wise covariance.
2. AAB-ALB-ND: spacing of apertures along branch and laterally as well as node size display moderately high pair-wise covariance.
3. BW-DW: branch width and dissepiment width display strong covariance.
4. AD-ND: aperture size and node size display moderately strong covariance.
5. Only the combination of aperture size (AD) and spacing of apertures along branch (AAB) displays a covariance notably lower than the rest of the pair-wise comparisons (Table 8).