| |
INTRODUCTION
The synonyms and homologies of muscular structures in the vertebrate head over their evolutionary history are highly debated resulting in several terminological confusions and controversies (Hoyos and Dubois 2004;
Diogo and Abdala 2010).
Dietrich Starck (1982, p. 7-8; own translation)
stated on the terminology of muscles: The nomenclature of singular muscles is not uniform and causes difficulties when
comparing different vertebrate classes. Primarily
adapted from human anatomy, non-comparable
muscles are assigned with the same name.
Additionally, many muscles were named by their
function without referring to the fact that no
muscle has only one function and that
homologous muscles in different taxa may have different
functions. To reach a more rational
nomenclature in Comparative Anatomy, it was attempted to
name muscles exclusively based on their origin
and insertion [...]." At this point he generally refers to the works of Max
Fürbringer and continues: "In addition, this approach had to fail, because origin and
insertion are not unchangeably fixed in phylogeny. Hence considerable discrepancies occurred.
Additionally, very unpractical sesquipedalians [word monsters] evolved, composed of up
to six or more singular terms. Currently, following
situation prevails [
]. For particular muscles
and muscles groups especially valuable for
primary musculature of the rump homologies
are clarified so far, hence, a consistent
nomenclature is possible for all vertebrate
classes.
 Easy and short designations are proved,
derivable from topographical relationships and
major courses. For mammals, the nomenclature
is designed after human anatomy, particularly
because homologies are widely verified. Apart
from that, it is strongly recommended to
transfer muscle names to other systematic taxa only if
evolutionary derivations are assessable.
Especially for musculature of the extremities of lower
tetrapods an independent nomenclature should
be used, because the adaptation of terms out of
mammal-anatomy pretends homologies, which are
not proved. Easy and short designations are proved,
derivable from topographical relationships and
major courses. For mammals, the nomenclature
is designed after human anatomy, particularly
because homologies are widely verified. Apart
from that, it is strongly recommended to
transfer muscle names to other systematic taxa only if
evolutionary derivations are assessable.
Especially for musculature of the extremities of lower
tetrapods an independent nomenclature should
be used, because the adaptation of terms out of
mammal-anatomy pretends homologies, which are
not proved.
In the present contribution, I first set clear definitions of such basic and crucial terms as origin, insertion, muscle portion, and muscle section in cranium-associated muscular structures (Figure 1). For the cranial musculature of turtles, I then introduce both a simple numeration system of muscular units and – partly polysyllabic – muscle names. Although criticised by
Starck (1982), the latter are unavoidable due to 1.
nomenclatural traditions and 2. muscular units – the smallest part of macroscopic muscular structures (see below) – have a high anatomical resolution. As such, they require explicit terms.
Starck (1982, p. 8) stated that regarding the m. adductor mandibulae structures: "One stable terminology is exemplarily designed for the n. trigeminus supporting jaw musculature. Despite various differences in particular the homologies for all non-mammal taxa is widely resolved. A uniform nomenclature is possible." As shown below, this statement is correct regarding the superficial similarity of those muscles, but the homology of detailed differentiations of muscle portions and muscle heads are not understood in detail yet. Major structures such as m. adductor mandibulae posterior (No. 29-30; for the introduced numeration of muscular units see below) are the subject of various discussions due to uncertain homologisations.
The anatomy of the turtle head is of particular interest due to the uncertain position of turtles within amniotes (Rieppel 2008;
Werneburg and Sánchez-Villagra 2009). Either turtles plesiomorphically show an anapsid skull or, when nested within diapsids, a secondary anapsid skull. In the latter case two temporal fenestra of the dermatocranium would have closed in evolution (Müller 2003).
 Jaw and cranium-associated neck musculature are assumed to have an important mechanical influence on the formation of dermatocranial bone arrangements in the temporal region of the skull (e.g.,
Frazetta 1968;
Smith 1996;
Rieppel 1993;
Tarsitano et al. 2001). Several studies have proposed homologies and evolutionary transformations of this musculature across reptilian groups (Holliday 2006;
Tvaroková 2006;
Holliday and Witmer 2007;
Diogo et al. 2008a,
2008b). Jaw and cranium-associated neck musculature are assumed to have an important mechanical influence on the formation of dermatocranial bone arrangements in the temporal region of the skull (e.g.,
Frazetta 1968;
Smith 1996;
Rieppel 1993;
Tarsitano et al. 2001). Several studies have proposed homologies and evolutionary transformations of this musculature across reptilian groups (Holliday 2006;
Tvaroková 2006;
Holliday and Witmer 2007;
Diogo et al. 2008a,
2008b).
Classic studies on the head muscle anatomy of single turtles species include those of Wiedemann (1803),
Bojanus (1819-21), and
Ogushi (1913b). More
comprehensive studies between different turtle taxa were published by
Hoffmann (1890),
Lakjer (1926),
Schumacher (1953-54,
1954,
1954-55a,
1954-55b,
1972), and
Poglayen-Neuwall (1953a).
Schumacher (1973) summarised the literature and set an often-cited standard when referring to head musculature in turtles (e.g.,
Wochesländer et al. 1999;
Wynecken 2003). From 313 known turtle species and 97
Wiedemann (1803),
Bojanus (1819-21), and
Ogushi (1913b). More
comprehensive studies between different turtle taxa were published by
Hoffmann (1890),
Lakjer (1926),
Schumacher (1953-54,
1954,
1954-55a,
1954-55b,
1972), and
Poglayen-Neuwall (1953a).
Schumacher (1973) summarised the literature and set an often-cited standard when referring to head musculature in turtles (e.g.,
Wochesländer et al. 1999;
Wynecken 2003). From 313 known turtle species and 97
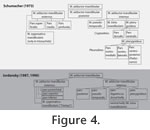 genera (sensu
Fritz and Havaš 2007), the jaw musculature of more than 40 species and more than 30 genera are described (e.g.,
Figure 2,
Figure 3,
Figure 4). In contrast, the anatomy of neck musculature has been described only for a few species, with a focus on general differences of the neck muscles between side-necked (Pleurodira) and hidden-necked (Cryptodira) turtles (Vallois 1920,
1922;
George and Shah 1955b;
Shah 1963;
Scanlon 1982;
Herrel et al. 2008). Other muscle systems of the head – external and internal ocular, laryngeal, and hyobranchial muscles – are described only for a few species (von Bayern 1884;
König 1934-35;
Gräper 1932,
1932;
Nishi 1916,
1938b;
Gaunt and Gans 1969;
Schumacher 1972). genera (sensu
Fritz and Havaš 2007), the jaw musculature of more than 40 species and more than 30 genera are described (e.g.,
Figure 2,
Figure 3,
Figure 4). In contrast, the anatomy of neck musculature has been described only for a few species, with a focus on general differences of the neck muscles between side-necked (Pleurodira) and hidden-necked (Cryptodira) turtles (Vallois 1920,
1922;
George and Shah 1955b;
Shah 1963;
Scanlon 1982;
Herrel et al. 2008). Other muscle systems of the head – external and internal ocular, laryngeal, and hyobranchial muscles – are described only for a few species (von Bayern 1884;
König 1934-35;
Gräper 1932,
1932;
Nishi 1916,
1938b;
Gaunt and Gans 1969;
Schumacher 1972).
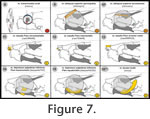
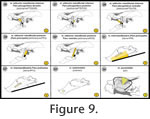
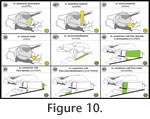
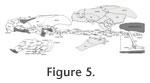 In the present study, I review all cranium-associated muscular structures that – to my knowledge – were ever described for turtles in the literature (Appendix 1,
Figure 5,
Figure 6,
Figure 7,
Figure 8,
Figure 9,
Figure 10,
Figure 11,
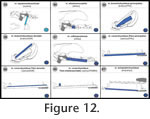
Figure 12,
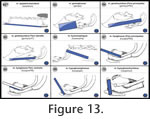
Figure 13,
Figure 14,
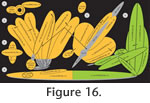
Figure 15,
Figure 16). In the present study, I review all cranium-associated muscular structures that – to my knowledge – were ever described for turtles in the literature (Appendix 1,
Figure 5,
Figure 6,
Figure 7,
Figure 8,
Figure 9,
Figure 10,
Figure 11,
Figure 12,
Figure 13,
Figure 14,
Figure 15,
Figure 16).
 The internal and external ocular muscles are innervated by n. oculomotorius (III) (No. 1-8), n. trochlearis (IV) (No. 9-10), and n. abducens (VI) (No. 34-38). The palatoquadrate, nose, and lower orbit region related musculature is innervated by n. trigeminus
(V). The jaw depressor muscle, the superficial neck musculature and muscular structures medial to the ear capsule are innervated by n. facialis (VII) (No. 39-46). The internal and external ocular muscles are innervated by n. oculomotorius (III) (No. 1-8), n. trochlearis (IV) (No. 9-10), and n. abducens (VI) (No. 34-38). The palatoquadrate, nose, and lower orbit region related musculature is innervated by n. trigeminus
(V). The jaw depressor muscle, the superficial neck musculature and muscular structures medial to the ear capsule are innervated by n. facialis (VII) (No. 39-46).
 N. glossopharyngeus (IX) innervates branchiovisceral musculature (No. 47-48). N. vagus (X) and n. accessorius (XI) support laryngeal (No. 49-51), as well as one n. vagus (X) innervated shoulder muscle (No. 52) and its derivate (No. 53). The muscles of the tongue, the hyoid apparatus, and a few muscles N. glossopharyngeus (IX) innervates branchiovisceral musculature (No. 47-48). N. vagus (X) and n. accessorius (XI) support laryngeal (No. 49-51), as well as one n. vagus (X) innervated shoulder muscle (No. 52) and its derivate (No. 53). The muscles of the tongue, the hyoid apparatus, and a few muscles
 of the neck region (No. 54-74) gain their innervation by n. hypoglossus (XII). Most neck-related muscles are exclusively innervated by spinal nerves, but depending on the anterior-posterior extend of the muscles as well as on their ontogenetic origin – e.g., from visceral mesoderm – they may also be partly innervated by posterior cranial nerves; or – if originating from postcervical myotoms – also by nerves of the trunk and tail region (No. 75-88). of the neck region (No. 54-74) gain their innervation by n. hypoglossus (XII). Most neck-related muscles are exclusively innervated by spinal nerves, but depending on the anterior-posterior extend of the muscles as well as on their ontogenetic origin – e.g., from visceral mesoderm – they may also be partly innervated by posterior cranial nerves; or – if originating from postcervical myotoms – also by nerves of the trunk and tail region (No. 75-88).
 As a case study to demonstrate the presented standardisation of cranium-associated musculature in turtles, I studied the cranial anatomy of the red-bellied short-necked turtle Emydura subglobosa (Krefft 1876). It is a common carnivore species living in freshwater environments of Northern Australia and Papua New Guinea (Legler and Georges 1993). Due to its easy keeping and breeding requirements (Nicol 1993; As a case study to demonstrate the presented standardisation of cranium-associated musculature in turtles, I studied the cranial anatomy of the red-bellied short-necked turtle Emydura subglobosa (Krefft 1876). It is a common carnivore species living in freshwater environments of Northern Australia and Papua New Guinea (Legler and Georges 1993). Due to its easy keeping and breeding requirements (Nicol 1993;
 Highfield 1996;
Hennig 2001;
Pawlowski 2001;
Schwarz 2006), E. subglobosa was proposed to be a suitable pleurodiran "model" species for evolutionary developmental studies (Tzika and Milinkovitch 2008;
Werneburg et al. 2009). By describing the adult head morphology of this species, I would like to expand knowledge of those structures in pleurodire turtles, which are underrepresented in most studies.
Kesteven (1942-1945) and
Schumacher (1954-55a) were the first to describe the cranial musculature of an Emydura-species, E. macquarii kreftii (Gray 1831), a species possibly opposing all remaining Emydura-species, and E. subglobosa is nested deep within the Emydura-clade (Georges and Thomson 2006;
Iverson et al. 2007). The development of the external morphology and ossification patterns of E. subglobosa were recently described (Werneburg and Sánchez-Villagra 2009;
Werneburg et al. 2009). Highfield 1996;
Hennig 2001;
Pawlowski 2001;
Schwarz 2006), E. subglobosa was proposed to be a suitable pleurodiran "model" species for evolutionary developmental studies (Tzika and Milinkovitch 2008;
Werneburg et al. 2009). By describing the adult head morphology of this species, I would like to expand knowledge of those structures in pleurodire turtles, which are underrepresented in most studies.
Kesteven (1942-1945) and
Schumacher (1954-55a) were the first to describe the cranial musculature of an Emydura-species, E. macquarii kreftii (Gray 1831), a species possibly opposing all remaining Emydura-species, and E. subglobosa is nested deep within the Emydura-clade (Georges and Thomson 2006;
Iverson et al. 2007). The development of the external morphology and ossification patterns of E. subglobosa were recently described (Werneburg and Sánchez-Villagra 2009;
Werneburg et al. 2009).
|